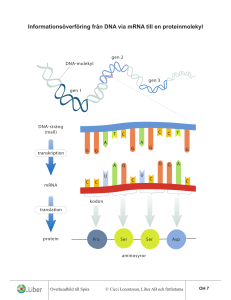
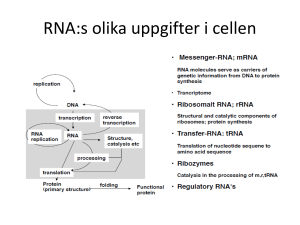
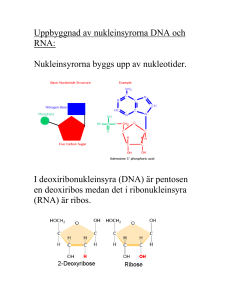
Transkriptionsinitiering / genreglering (FH)
advertisement
")
Transkriptionsinitiering / genreglering (FH) Skillnad mellan pro- och eukaryot bildning av av mRNAt: Prokaryot genreglering: I många fall regleras gener tillsammans i sk Operon (ex. Lac- eller trp-operonet) Lac operonet: Exempel på reglerat genuttryck med hjälp av en sk. repressor. Lac-repressorn är trans-verkande dvs det är inte nödvändigt att genen för Lacrepressorn sitter på samma DNA sträng som den gen eller regleringselement som repressor proteinet verkar på. Motsatsen till trans- är cis-verkande. För cis-verkande element är det nödvändigt att genen/elementet sitter på samma DNAsträng. Promotorn: Styr transkriptions initiering - RNA polymeraset binder till promotorn Prokaryota promotorner har två konserverade delar (-10 (TATAAT) samt -35 (GACAT) regionerna, sk. boxar. +1 definieras av den nukleotid som motsvarar den första nukleotiden i RNAt. RNA polymeraset är ett komplex av flera proteinsubenheter med grundstrukturen (2α2βσ) 2α enheter samt 2β enheter. Dessutom finns det en σ enhet (faktor). σ - faktorn har betydelse vid initieringen och den binder till TATAA boxen i promotorn. Eukaryot genreglering: Man pratar ofta om genexpression eller genuttryck, med detta menas vilka gener som är ”påslagna” eller ”aktiva” och som producerar en produkt i en individ eller en specifik cell och bidrar till fenotypen. Expressionen/uttrycket av en proteinkodande gen kan delas upp i mRNA-uttryck och proteinuttryck och kan mätas genom att mäta hur mycket mRNA resp. protein det finns i tex celltypen ("nivån" av tex ett specifikt mRNA). mRNA expressionen är en produkt av hur aktiv en gen är (hur många mRNA som produceras per tidsenhet) och dels hur stabilt ett mRNA är. mRNA nivå = resultatet av syntesen (hur aktiv en gen är) och nedbrytning (hur stabilt ett mRNA är) 1 Reglering av mRNA expressionen sker på olika sätt: Den större genetiska komplexiteten hos eukaryoter (fler gener som dessutom är enskilt reglerade) än hos prokaryoter, kräver en mer omfattande kontrollapparat. Reglering av transkriptionsinitieringen är det enskilt viktigaste regleringssättet men det finns andra: 1. Kromatinstrukturen påverkar hur tillgänglig en gen är för transkription: Reglerad acetylering av histoner eller metylering av DNA 2. Transkriptionsinitiering Genaktivitet. Hur ofta det görs ett mRNA 3. mRNA stabilitet Man pratar om att ett mRNA har en viss livsläng eller halveringstid (poly Asvansens längd samt förekomst av de-/stabiliserand sekvenser i mRNAt 5’ och 3’ UTR påverkar stabiliteten). 4. Komplexa transkriptionsenheter Alternativ splitsning, resp. alternativ polyadenylering Transkriptionsreglering: Det finns oftast flera olika regulatoriska element som styr en gens aktivitet. Promotorn Andra element (cis-acting) som aktiverar eller inhiberar genaktivitet. Promotor proximala element, enhancers, silencers Det finns 3 (4) olika eukaryota RNA polymeras med olika uppgifter: RNA polymeras-I: syntetiserar rRNA (pre-rRNA =>28S, 5.8S, 18S rRNA) -II: syntetiserar mRNA samt 4 st snRNA (sk. URNA) -III: syntetiserar tRNA, 5S rRNA samt div. sn- och snoRNA samt (mitokondrie/kloroplast RNA polymeraser) De 3 eukaryota RNA polymerasen har en liknande grundstruktur som det prokaryota RNA polymeraset; 2αββ', men saknar σ-faktorn. mitokondrie/kloroplast RNA polymeraser skiljer sig. Promotorns struktur/sekvens: Varierar mycket mer mellan olika gener hos eukaryoter än hos prokaryoter. Core-promotor En del eukaryota promotorer har en sk. TATA-box vid position –34, andra en sk initiator-sekvens direkt vid +1. Promotorns styrka: bestämmer hur ofta ett RNA pol initierar transkription från promotorn. En ”stark” promotor framför en gen leder till hög genaktivitet. 2 Transkriptionsinitiering proteinkodande gener: (gäller RNA pol II): RNA pol II bildar tillsammans med andra proteiner ett sk. transkriptionspreinitieringskomplex (preinitieringskomplex). Dessa andra proteiner kallas generella transkriptionsfaktorer (TF). Transkriptionsfaktorerna (TFs) i initieringskomplexet behövs för att aktivera RNA pol II samt att binda till proteinkodande geners promotorer. De finns i alla typer av celler, därav namnet generella transkriptionsfaktorer. Bland de generella transkriptionsfaktorerna finns TATA-box binding factor (TBP), faktorer med kinasaktivitet och som kinaserar CTD på RNA pol II (TFIIH) Bildandet av initieringskomplexet är nödvändigt för att transkription ska ske men det är ofta inte tillräckligt. Aktivatorer / Specifika TFs För att transkriptionen skall ske krävs fler transkriptionsfaktorer. Dessa mer specifika TF finns i olika uppsättning tex. i nervceller eller muskelceller och kallas; specifika TFs, eller vävnadsspecifika TFs. Det är dessa faktorer som bestämmer vilka gener som kommer att vara aktiva i olika vävnader. Specifika TFs binder till regulativa element Specifika TFs är oftast aktiverande men kan också inhibera transkription, och kallas då repressorer. Specifika TFs kan binda till DNA med specifik sekvens. Specifika TF binder DNA antingen till reglerings-element i eller strax uppströms om promotorn eller till element som kan sitta långt från promotorn. Reglerings-element som ej sitter i direkt anslutning till promotorn och medierar aktivering kallas "enhancers" och element som medierar inhibering kallas "silencers". Co-aktivatorer och mediatorer vid direkt interaktion mellan initiering komplex och TF Både bildandet av preinitieringskomplexet och bindningen till promotorn underlättas av sk. co-aktivatorer (TranskriptionsAktiverande Faktorer, TAFs eller sk. ”mediatorer”). Co-aktivatorer sitter ofta ihop i ett komplex. Co-aktivatorerna förmedlar i dessa fall en direkt kontakt mellan specifika TFs och preinitieringkomplexet så att transkription kan starta vid promotorn. Det finns också co-repressorer och komplex av dessa som verkar på liknande sätt men motsattverkan. Indirekt interaktion via Kromtinstruktur Vissa specifika TFs och co-faktorer kan påverka kromatinstrukturen så att den packas upp (dekondenseras) eller tätpackas (kondenserat kromatin) och kan på detta indirekta sätt påverka genaktiviteten hos gener som ligger i regionen. Kromatin kan dekondenseras bl.a. genom att histoner acetyleras och detta kan ske genom att specifika TFs binder till ”enhancer” element och ”tar med sig” histonacetylaser till området. Promotorn görs tillgänglig för transkriptionsinitiering och hela genen kan transkriberas. Betydelsen av histonacetylering och methylering samt hur TFs binder till ex enhancers och drar till sig acetyl transferas, samt motsatsen då de-acetylaser samt methylaser lockas till regionen med TFs. 3 Betydelsen vid methylaser och mer långsigtig inhibering av regioner – betydelsen av methyl histon bindande proteiner samt proteiner som i sin tur lockas till regionen och kondenserar kromatinet. Lodish – bra. TFs har DNAbindande domän Karakteristiskt för många specifika TF är att de är aktiva som dimerer och kan binda till DNA med hjälp av en sk. DNA-bindande domän. De har ofta ytterligare domäner som kan vara tex. protein-bindande. Den DNA bindande domänen kan ha olika protein domänmotiv (finns ett 10-tal olika) Ex.: Zink-fingermotiv (ett av de vanligaste motiven) Homeodomän ("Helix turn Helix "motiv) "basic Helix-loop-Helix" motiv "Leucine-zipper" motiv OBS kan tas på genexpressionsanalys föreläsningen! De specifika TF medierar vävnadsspecifikt genuttryck. Beroende på vilken sekvens promotorn har och vilka reglerings-element som är kopplade till genen så aktiveras olika gener av olika specifika TF. Uppsättningen av specifika TF bestämmer alltså vilka gener som kan vara aktiva i en specifik celltyp eller vävnad. Detta kallas vävnadsspecificitet. Reglering/Inducering av genaktivitet För att genen skall vara aktiv krävs att alla komponenter som aktiverar initieringen finns på plats! Reglering av genaktivitet kan därför ske genom att avlägsna eller lägga till nödvändiga specifika TF. Detta sker genom att tex flytta dem ut och in i kärnan eller att påverka deras förmåga att binda DNA eller protein Två exempel på hur detta kan ske: Ex. 1. Nuklära receptorer är både receptorer och specifika TF. De kan först efter det att de bundit en specifik ligand fungera som en TF. På detta sätt fungerar många av våra steroid hormon (tex. östrogen). Efter ligandbindning så aktiverar de transkriptionen av de specifika gener till vars regleringselement de kan binda. Ex. 2. Aktivering av TF Stat1α: Om Stat1 fosforyleras av ett proteinkinas (JAK) så kan det bilda dimerer och förs därmed in i kärnan och kan aktivera transkription av de specifika gener där Stat1 kan binda. Fosforylering -> transport till kärnan från cytoplasman. 4