Kap 7 och 8 Genreglering
advertisement

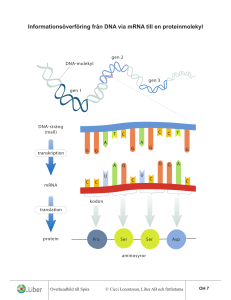
Kontroll av genuttrycket på transkriptionsnivå 7.1-7.6 Vägen från gen till funktionellt protein består av många steg, fig 1-11. Det finns exempel på kontrollmekanismer för alla dessa steg. Transkriptionsstart Alternativ splicing Nedbrytning av mRNA Translationsstart Proteinets stabilitet, aktivitet och lokalisering Synteshastigheten för ett protein avgörs av hur ofta ………………………. av genen och ……………………………. av mRNA´t sker. Hur ofta en gen transkriberas beror på hur bra ……………………………… binder till promotorn. Det sker ingen (eller väldigt sällan) bindning om DNA´t är i ………………………..-kromatinstruktur . Olika promotorsekvenser har olika hög affinitet för RNA-polymeraset. Mha ……………………………………….. kan affiniteten både ökas och minskas. Mha en …………………….får RNA-polymeraset ytterligare en bindningsmöjlighet till DNA´t (förutom promotorn), därmed ökas affiniteten. En ………………………… stör bindningen mellan RNA-polymeras och DNA. Reglering av Lac-operonet är ett bra exempel på hur en repressor och en aktivator kan fungera i en bakterie. Lac-operonet kodar för proteiner som behövs för att cellen ska kunna ta upp och klyva disackariden laktos. Laktos glukos + galaktos. Detta måste ske för att cellen ska kunna använda laktos som energikälla. Om cellen inte har tillgång till laktos är lac-operonet avstängt. Transkription sker väldigt sällan. Om cellen har tillgång till laktos men också glukos sker transkription, men med ganska låg frekvens. Cellen sparar energi genom att använda glukos isf laktos som energikälla. Om cellen bara har tillgång till laktos och inte glukos uttrycks lacoperonet med hög frekvens. Till lac-operonet hör de reglerande sekvenserna: promotor,………………………...och …………………..., fig 7-2. Utan laktos binder lac repressorn till operatorn och hindrar då polymeraset från att binda till promotorn (7-2a). Med laktos lossnar repressorn och ………………………. kan binda promotorn, men bindningsaffiniteten är låg (7-2b). Med laktos utan glukos binder cAMP till CAP-site och till polymeraset som då får en högre affiniteten till DNA´t (7-2c). Transkription hos eukaryoter skiljer sig på några punkter från prokaryoter: • Eukaryoter har tre olika RNA-polymeraser (I, II och III), tabell 7-2. • Det finns inga eukaryota sigmafaktorer, i stället behövs sk ………………………. transkriptions-faktorer för att binda polymeraset till promotorn. • Det krävs många steg innan transkriptionen kan börja, DNA´t skall vara i form av eukromatin och ett antal specifika och allmänna transkriptionsfaktorer skall binda DNA och polymeras. RNA-polymeras II´s promotor-sekvens kan innehålla sk TATA-box (fig 7-12), initiator-sekvens eller områden med mycket CG-baspar. För att RNA-polymeraset ska kunna bindas till promotorn och transkriptionen starta måste allmänna transkriptionsfaktorer medverka. RNA-polymeras II har hjälp av TFIIA, B osv., fig 7-31. (TF=transcription factor) Eukaryota gener har förutom promotorsekvenser ett flertal andra reglerande sekvenser, fig 7-16. En del finns nära genen, inom några hundra bp. Andra kan finns mycket långt bort, 10.000-tals bp. Dessa kallas ……………….-sekvenser eftersom det ofta är aktivatorer som binds dit. Till dessa reglerande sekvenser binds …………………………………………………………… Specifika transkriptionsfaktorer har en DNA-bindande domän och en eller flera aktiverande/inhiberande domäner, fig 7-22. I kapitel 16 kommer exempel på en massa olika transkriptionsfaktorer, se t ex fig 16-1. I transkriptionsfaktorns DNA-bindande domän finns något DNAbindande motiv. Ofta är det en ............ som binds till major groove i DNA´t. Exempelvis i helix-turn-helix, leucin zipper och zink-fingrar, fig 724 --- 7-26. Helix-turn-helix Leucin-zipper Ofta består de reglerande sekvenserna i DNA´t av två proteinbindande sekvenser bredvid varandra. Det ger möjlighet att kombinera olika specifika transkriptionsfaktorer med varandra till ett större antal ”signaler”. Tre olika proteiner kan t ex kombineras på sex olika sätt, fig 7-28. De ………………..transkriptionsfaktorerna är desamma för olika gener medan kombinationen av de …………..……………. är unik för varje gen. En gen kan fås att uttryckas i en viss vävnad genom att ha reglerande sekvenser som binder aktivatorer som bara finns i den vävnaden. Ex: genen för proteinet transtyretin, fig 7-44. Transkriptionsfaktorer och RNA-polymeras binder lättare till DNA´t om detta är i eukromatin-form. Gener kan tystas genom att DNA´t övergår till heterokromatin. Heterokromatin kan bildas genom att histonsvansar…………………………….. Vid replikation kan histonernas modifieringsmönster ”ärvas” till båda dotterkromosomerna. Detta kallas epigenetik. Fig 7-36 visar ett exempel på epigenetik. Hox-gener tystas tidigt under embryogenesen av en och förblir avstängda livet ut. Vid replikationen bildas nya histonproteiner som aggregerar till sk nukleosomkärnor, dvs 2x(H2A, H2B, H3, H4). De gamla nukleosomkärnorna fördelar sig på båda de nya kromosomerna. Om de är metylerade sprids metylering från dem till de nygjorda histonerna. Me Me Me Me “Gammal” nukleosomkärna Ny nukleosomkärna Aktivering av en eukaryot gen skulle kunna gå till så här: 1) Till en reglerande sekvens binds en transkriptionsfaktor som i sin tur binder ett enzym som acetylerar histonsvansar i närliggande nukleosomer, fig 738b. Det leder till att fler reglerande sekvenser exponeras. 2) Då kan specifika transkriptionsfaktorer, mediatorer, allmänna transkriptionsfaktorer och RNA-polymeras bindas samman så att transkriptionen kan starta, fig. 7-42 RNAi, sid 347-351 Det finns RNA´n som har till uppgift att reglera genuttrycket, sk interferensRNAn eller RNAi. Många viktiga processer verkar regleras av RNAi, bl a fosterutvecklingen, fig 8-27. RNA-interferens är ett mycket aktivt forskningsfält. Man skulle t ex kunna skräddarsy gener för mi/siRNA som är riktade mot ett specifikt mRNA man önskar slå ut. Det finns två typer av RNAi: miRNA (microRN) och siRNA (short interferenceRNA). Ex på hur ett miRNA fungerar (ungefär samma sak sker med siRNA), fig 8-26: Människan har minst 1000 gener för olika miRNAn. Det primära transkriptet kan var flera 100 nt långt. I kärnan viker det ihop sig, binds till proteiner (bl. a Drosha) och trimmas till en kortare sekvens. miRNA´t transporteras ut ur kärnan. Proteinkomplexet med Drosha blir kvar i kärnan. I cytosolen binds miRNA´t till ett nytt proteinkomplex (bl.a Dicer). miRNA´t klyvs så att två korta, 21-23 nt, strängar bildas. Den ena av de två korta strängarna binds till ett nytt proteinkomplex. Då bildas ett sk RISCkomplex (RNAinduced silencing complex). Det är det reglerande komplexet. Ett RISC-komplex med miRNA binder till ett mRNA som har en nästan komplementär sekvens till miRNA´t, fig 8-25. Resultatet blir att translationen av mRNA´t förhindras. Ett RISC-komplex med siRNA binder till ett mRNA som har en exakt komplementär sekvens till siRNA´t. Resultatet blir att mRNA´t bryts ner. RISC-komplexet fungerar då som ett nukleas.