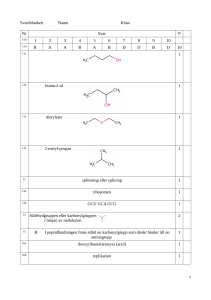
a) Från värdena på olika G-parameterar kan man få värdefull
advertisement
 Från värdena på olika G-parameterar kan man få värdefull")
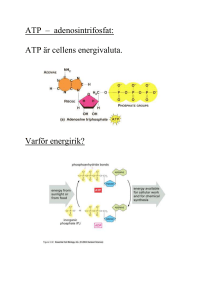
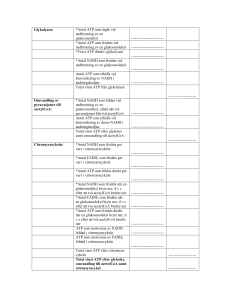
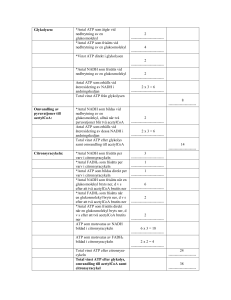
LINKÖPINGS UNIVERSITET IFM/Kemi TENTAMEN I BIOKEMI 1 NKEA08, 92KE21, 92KE27, 9KE321, 9KE361 2012-12-21 kl. 08.00-12.00 Skriv endast en uppgift per blad. Redovisa beräkningar och motivera svar. Besvara delfrågor i tur och ordning. Hjälpmedel: miniräknare Ansvariga lärare: Magdalena Svensson (285686, 0704-090999) och Anki Brorsson (286648, 0762-209433) 1.a) Rita följande tetrapeptid: Phe-Val-Met-Asn vid pH 7. Rita/markera huvudkedja, sidokedjor, α-kol, N- och C-terminal. Ange även vilken nettoladdning tetrapeptiden har vid pH 7. (4p) b) Rita två H-bindningar; en som är en del av sekundärstrukturen och en annan som är en del av tertiärstrukturen. (4p) c) Sekundärstruktur i proteiner består av α–helixar och β–flak. Ange två skillnader mellan dessa strukturtyper. (2p) 2.a) Med avseende på vad sker separationen av proteiner i följande tekniker? i) gelfiltreringskromatografi ii) affinitetskromatografi iii) jonbyteskromatografi (3p) b) Vilken är den principiella skillnaden mellan PAGE och SDS-PAGE samt vilka konsekvenser får det för din analys av resultatet? (2p) c) Du har en peptid, Ala-Trp-Ser-Phe-Lys-Met-Glu-Arg-Gly, till vilken du tillsätter trypsin. Vad blir resultatet? (2p) d) Du funderar nu på att göra några olika mutanter av trypsin (ett vattenlösligt globulärt protein). i) I trypsins aktiva yta finns en Ser. Vilken aminosyra bör du byta till om du vill ha en så liten påverkan av proteinaktiviteten som möjligt? Motivera! ii) På ytan av trypsin finns en cis-Pro. Vilken aminosyra bör du byta till om du vill ha en så stor påverkan av proteinaktiviteten som möjligt? Motivera! iii) I det inre av trypsins struktur finns en Leu. Vilken aminosyra bör du byta till för att det ska blir en så liten påverkan som möjligt av proteinaktiviteten? (3p) 3. a) En enzymkatalyserad reaktion påverkas bland annat av temperaturen. Beskriv, schematiskt, hur du skulle kunna gå till väga för att bestämma den optimala temperaturen för ett visst enzym (dvs den temperatur där enzymet uppvisar högst aktivivitet). (3p) b) Förklara följande två begrepp: i) zymogenaktivering ii) allosterisk kontroll (4p) c) ATP-syntas är ett multienzymkomplex som katalyserar ATP-syntes. i) Var i cellen sker syntesen? ii) En sidokedja till Asp hos ATP-syntas spelar en viktig roll, vilken? (3p) 4. a) Glykolysens slutprodukt är pyruvat. Vad kan ske med pyruvat under anaeroba förhållanden och hur mycket ATP bildas då en glukosmolekyl bryts ner under anaeroba förhållanden? (2p) b) Omvandlingen från fosfoenolpyruvat till pyruvat är det sista steget i glykolysen och den omvända reaktionen är inledningen på glukoneogenesen. Beskriv vad som skiljer dessa reaktioner åt och varför detta är viktigt. (3p) c) Nämn två metaboliter som kan fungera som ingångsämnen för glukoneogenesen. (1p) d) Pyruvatdehydrogenaskomplexet är ett viktigt kontrollenzym för citronsyracykeln. Vilken reaktion katalyseras av pyruvatdehydrogenaskomplexet? (1p) e) Hur påverkas aktiviteten hos pyruvatdehydrogenaskomplexet om koncentrationerna nedan gäller? (motivera ditt svar) (2p) [NADH] > [NAD+] [AcetylCoA] > [CoA] [ATP] > [ADP] f) Vilka två viktiga syften har citronsyracykeln? (1p) 5. Nedbrytning och syntes av fettsyror regleras hormonellt av glukagon och insulin genom kovalent modifiering av enzymerna acetylCoAkarboxylas och lipas a) Vilka reaktioner katalyseras av dessa enzymer? (2p) b) Vilken form av kovalent modifiering är det som sker? (1p) c) Hur regleras enzymerna av respektive hormon (aktiveras eller inhiberas)(motivera)? (4p) d) I en hälsokostaffär såldes chokladbitar med tillsats av karnitin. Ge en biokemisk motivering till varför tillverkaren anser att det är vettigt att tillsätta karnitin i chokladen. (Om karnitin administrerad på detta sätt verkligen har någon effekt förblir osagt). (2p) 6. Följande steg sker vid fettsyranedbrytningen Oxidation Hydratisering Oxidation Klyvning a) I vilken sorts reaktion genereras NADH och FADH2 (1p) b) Via andningskedjan genereras ATP från NADH och FADH2. Varför erhålls mer ATP från NADH jämfört med FADH2 (4p) c) Varför fås en mindre mängd ATP vid nedbrytning av omättade fettsyror? (1p) d) DNA och RNA är uppbyggda av nukleotider som är kopplade till varandra via en 3,5’ fosfodiesterbindning. Vad står 3’ och 5’ för i en 3,5’-fosfodiesterbindning? (1p) e) Vad innebär att replikationen är semikonservativ? (1p) f) Vilka är de vanligaste typerna av RNA och vilka funktioner har de? (3p) Lösningsförslag NKEA08, 92KE21, 92KE27, 9KE321 & 9KE361: 1. a) Nettoladdning = 0 b) Sekundärstruktur stabiliseras av H-bindningar mellan peptidgrupper, dvs NH och COO. Tertiärstruktur stabiliserar bland annat av H-bindningar mellan sidokedjor tex. Ser-OH och Asn-CONH2. c) Alfa-helix, H-bindningar parallellt med helix-axeln mellan aa relativt nära varandra i primärstrukture. Beta-flak, kan vara parallella eller antiparallella avseende kedjans riktning. H-bindningar vinkelrät mot kedjans riktning mellan aa vanlige rel. långt ifrån varandra i primärstrukturen. 2. a) i) storlek, ii) inbindningsförmåga till specifik ligand, iii) laddning b) Separation med PAGE sker efter storlek, form och laddning. Ett band på gelen anger att proteinet är rent (eller inte troligt men, ett protein med långsträckt form och hög laddning kan vandra lika långt som ett sfäriskt protein med låg laddning t.ex). Då vandringslängden beror av flera egenskaper hos proteinet går det inte (enkelt) att bestämma Mw för proteinet. Separation med SDS-PAGE sker efter storlek, minst vandrar längst. SDS är en negativt laddad detergent som binder in till polypeptidkedjan. Ett band på en SDSPAGE gel anger att proteinet är rent (eller att det är flera proteiner men med samma storlek). Vandringlängden för ett visst protein är omvänt proportionell mot lg Mw. Med hjälp av referensproteiner med kända Mw kan Mw för ett ”okänt” protein bestämmas. c) Trypsin klyber peptidbindningar C-terminalt om Arg och Lys.Resultat: Ala-TrpSer-Phe-Lys, Met-Glu-Arg, Gly. d) i) Aminosyror direkt inblandade i katalys svåra att byta ut. Men byte mot liknande kan fungera, t.ex Thr. (om man kan mekanismen för trypsin så är Cys den aa man väljer då Cys lätt att deprotonisera). ii) cis-Pro går i princip inte att byta utan stor påverkan, då för alla övriga aa dominerar trans-konfig med förhållandet 1000:1 mot cis. Så- vad vi än byter till får vi en stor effekt. iii. Leu i det inre av proteinet byt mot liknande sidokedja, opolär och alifatisk t.ex Ile. 3. a) Använd produkt med absorbans så reaktionen enkelt kan följas spektrofotometeriskt. Ha ett stort överskott av substrat så att tillgång på substrat inte begränsar reaktionshastigheten. Mät enxymaktivitet vid gradvis ökande temp. Mät produktbildning över tiden. Enzymaktivitet ökar med ökande temp dock vid för hög temp denaturerar enzymet. b) i) zymogenaktivering: ett zymogen (en inaktiv enzymform) aktiveras genom klyvning av den egna polypeptidkedjan. ii) allosterisk kontroll( produktinhibering spec.fall av allosterisk kontroll): allostera enzym ändrar sin konformation och därmed funktion då någon regleringsmolekyl binder in till regleringssite. c)i. ATP-syntas lokliserat till innermitokondriemembranet- själva syntesen sker i matrix ii) Det är denna Asp som möjliggör transport av protoner över innermitokondriemembranet vilket driver ATP.syntesen. Protoner binder till deprotoniserad Asp-sidokedja i c subenheterna (finns 10 st i cirkel). Till c kommer de genom en halvkanal i a. c-ringen roterar ett hack. Protoner lämnar c genom halvkanal i a. Transporten sker från innermitokondriutrymme till matrix. 4. a) I jästceller omvandlas pyruvat till etanol I muskelceller omvandlas pyruvat till laktat Energivinsten blir 2 ATP i båda fallen b) Fosfoenolpyruvat omvandlas till pyruvat i ett steg i glykolysen. Omvandlingen av pyruvat till fosfoenolpyruvat sker i två steg i glukoneogenesen. I första steget karboxyleras pyruvat till oxaloacetat och i andra steget dekarboxyleras och fosforyleras oxaloacetat till fosfoenolpyruvat. Det är viktigt att reaktionerna är olika så att de kan kontrolleras var för sig. Detta innebär att glykolysen kan kontrolleras separat från glukoneogenesen. När den ena processen är aktiv kan den andra stängas av. c) Laktat, aminosyror och glycerol (nämn två av dessa) d) Pyruvatdehydrogenaskomplexet katalyserar omvandlingen av pyruvat till acetylCoA (räcker som svar): Pyruvat + CoA + NAD+ → acetylCoA + CO2 + NADH + H+ e) Aktiviteten kommer att inhibiteras. Hög koncentration av ATP visar att det finns gott om energi och att citronsyracyken kan stanna av. Hög koncentration av acetylCoA och NADH innebär att det finns mycket produkt och att det inte behöver syntetiseras mer. f) Att generera energi (ATP) via reduktion av NAD+ och FAD till NADH och FADH2 som återoxideras i andningskedjan vilket resulterar i produktion av ATP. Att förse cellen med byggstenar som kan användas för att bilda andra molekyler som aminosyror, fettsyror och nukleotider mm 5. a) AcetylCoAkarboxylas katalyserar karboxylering av acetylCoA till malonylCoA Lipas katalyserar hydrolys av triacylglycerol till glycerol + fettsyror b) Fosforylering c) Glukagon fosforylerar lipas och gör det aktivt. Glukagon är ett hormon som produceras då vi behöver energi därför aktiveras fettsyranedbrytningen. Insulin defosforylerar lipas och gör det inaktivt. Insulin utsöndras efter måltid då kroppen har gott om energi och då inhiberas fettsyranedbrytningen eftersom det redan finns tillräckligt med energi. Glukagon fosforylerar acetylCoAkarboxylas och gör det inaktivt. Eftersom glukagon produceras när vi behöver energi så stoppas fettsyrasyntesen då det inte finns energi för att göra nya fettsyror. Insulin defosforylerar acetylCoAkarboxylas och gör det aktivt. Insulin produceras då det finns gott om energi och då aktiveras fettsyrasyntesen för produktion av nya fettsyror för att lagra energi. d) Karnitin behövs för att transportera in fettsyrorna till matrix i mitokondrien för vidare fettsyranedbrytning. Försäljningsargumentet är alltså att öka fettförbränningen. 6. a) NADH och FADH2 genereras i oxidationsstegen b) ATP bildas när H+ vandrar från mellanmembranutrymmet till matrix i mitokondrien via ATP syntas. En hög koncentration av H+ fås i mellanmembranutrymmet när elektroner från NADH och FADH2 vandrar genom andningskedjan. När elektronerna från NADH vandrar genom andningskedjan pumpas H+ från matrix till mellanmembranutrymmet vid tre proteinkomplex medan elektronerna från FADH2 bara passerar två. Elektronerna från NADH bidrar alltså mer till uppbyggnad av protongradienten över mitokondriens innermembran. c) Färre oxidationssteg vid nedbrytning av en omättad fettsyra leder till att det bildas en mindre mängd FADH2. d) En fosfatgrupp kopplar två sockerenheter via ett syre på 3’-kolet och ett syre på 5’kolet där 3’betyder kolatom nummer 3 i sockerenheten och 5’ betyder kolatom nummer 5 i sockerenheten. e) Efter replikationen kommer det i varje ny DNA-molekyl att finnas en ny och en gammal DNA-kedja. f) mRNA: Mall för proteinsyntesen. Bildas vid transkriptionen och används i translationen. rRNA: Beståndsdel i ribosomer. På ribosomerna sker translationen. tRNA: Hämtar aminosyror i cytosolen och bär dem till ribosomen. Ser till att rätt aminosyra hamnar på rätt plats i den växande polypeptidkedjan. Förutsättning för translationen. t