fettsyrametabolismen
advertisement

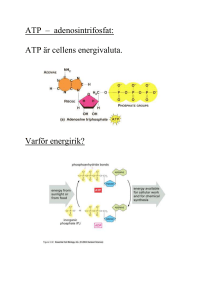
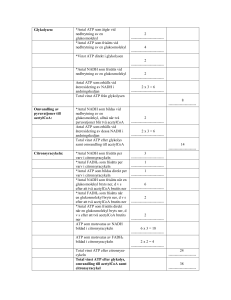
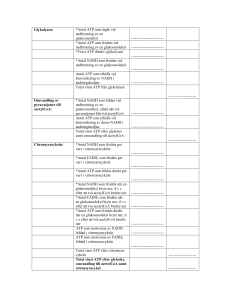
KOLHYDRATMETABOLISMEN: 1. Hur mycket ATP frisätts vid glykolysen och vad sker med pyruvat (i) vid aeroba förhållanden? (ii) anaeroba förhållanden? Svar: 2 ATP (i) Pyruvat bryts ner till CO2 + H2O i citronsyracykeln. (ii) Pyruvat kan i musklerna omvandlas till laktat och i mikroorganismer till etanol. 2. Hur skiljer sig glykolysen och glukoneogenesen åt på detaljnivå och vad är det som styr om respektive process är aktiv? Svar: Tre viktiga steg katalyseras av olika enzymer: * Pyruvat Fosfoenolpyruvat I glykolysen: Sker i ett steg och katalyseras av pyruvatkinas I glukoneogenesen: Sker i två steg; Pyruvat oxaloacetat katalyseras av pyruvatkarboxylas Oxaloacetat fosfoenolpyruvat katalyseras av fosfoenolpyruvatkarboxylas * Fruktos-1,6-bisfosfat fruktos-6-fosfat I glykolysen: Katalyseras av fosfofruktokinas I glukoneogenesen: Katalyseras av fruktos-1,6-bisfosfatas * Glukos-6-fosfat Glukos I glykolysen: Katalyseras av hexokinas I glukoneogenesen: Katalyseras av glukos-6-fosfatas Vilken process som är aktiv styrs främst av energibehov - behöver cellen energi är glykolysen aktiv och är det bättre att lagra energi till ett senare tillfälle är glukoneogenesen aktiv. 3. Vad innebär substratnivåfosforylering? Svar: Energi i form av en fosfatgrupp överförs direkt från en molekyl till ADP varvid ATP bildas. 4. Vilken reaktion binder samman glykolysen och citronsyracykeln? Svar: Pyruvats omvandling till acetylCoA. Pyruvat + CoA + NAD+ acetylCoA + CO2 + NADH + H+ Reaktionen katalyseras av pyruvatdehydrogenaskomplexet. 5. Redogör för hur mycket GTP, NADH och FADH2 som bildas per varv i citronsyracykeln. Svar: 1 GTP, 1 FADH2, 3 NADH 6. Vilka reaktioner gör citronsyracykeln energetiskt fördelaktig? Svar: De vars G0’ är negativt, främst AcetylCoA + oxaloacetat citrat Isocitrat -ketoglutarat + CO2 -ketoglutarat succinylCoA + CO2 7. Ge exempel på andra biomolekyler som kan bildas från citronsyracykelintermediärer. Svar: Från oxaloacetat och -ketoglutarat kan aminosyror och nukleotider bildas, från succinylCoA kan hem bildas som är en prostetisk grupp bl a i hemoglobin och nedbrytning av citrat kan ge acetylCoA som är utgångsämne för fettsyrasyntesen. 8. Beskriv översiktligt hur e- från NADH vandrar mot den slutliga e--acceptorn O2. Svar: Elektroner lämnas vid NADH-Q-oxidoreduktas (komplex I) och vandrar sedan via Q (ubiquinon) till Qcytokrom c oxidoreduktas (komplex III). Därifrån bärs elektronerna av cytokrom c till cytokrom c oxidas (komplex IV) för att slutligen lämnas till O2. 9. Förklara varför oxidation av NADH motsvarar 2.5 ATP medan oxidation av FADH2 motsvarar 1.5 ATP. Svar: När elektronerna från NADH vandrar genom andningskedjan pumpas H+ från matrix till inter- membransutrymmet vid tre proteinkomplex medan elektronerna från FADH2 bara passerar två. Elektronerna från NADH bidrar alltså mer till uppbyggnad av protongradienten. 10. Vad gör cytokromer till bra elektronbärare? Svar: Cytokromerna har en hemgrupp som prostetisk grupp och i denna finns en central järnjon som kan alternera mellan Fe2+ och Fe3+. FETTSYRAMETABOLISMEN: 11. Ange korrekt svarsalternativ (a eller b): (i) Fettsyrasyntesen sker a. i mitokondrien. b. i cytosolen. (ii) Fettsyrasyntesen a. kräver ATP. b. genererar ATP. (iii) Fettsyranedbrytningen katalyseras av a. flera separata enzymer. b. ett enzymkomplex. (iv) Vid fettsyrasyntesen bildas fettsyran a. från metyländen till karboxyländen. b. från karboxyländen till metyländen. (v) Lipas katalyserar a. bildandet av fettsyror. b. nedbrytning av triacylglycerol. (vi) Lipasaktiviteten regleras a. hormonellt. b. av allostera effektorer. (vii) Enzymet som reglerar hastigheten för fettsyrasyntesen heter a. acetylCoAdehydrogenas b. acetylCoAkarboxylas (viii) Insulin Svar: b, a, a. aktiverar fettsyrasyntesen. b. inaktiverar fettsyrasyntesen. a, a, b, a, b, a 12. Ange om det behövs oxidationsmedel eller reduktionsmedel för fettsyranedbrytningen och ange vilket/vilka. Svar: För nedbrytningen krävs oxidationsmedlena NAD+ och FAD. 13. Rita upp fettsyran 18:29,12 (linolsyra). Svar: CH3CH2CH2CH2CH2CH=CHCH2CH=CHCH2CH2CH2CH2CH2 CH2CH2COOH 14. Varför omvandlas acetylCoA till malonylCoA innan fettsyrasyntesen och det mekanistiska motivet dekarboxylering återfinns även i en andra metabola processer – vilka? Svar: Dekarboxylering sker när acetyl-ACP kondenserar med malonyl-ACP. Det byggs in energi i malonyl-ACP eftersom ATP åtgår. Att slå ihop två acetyl-ACP är energetiskt ofördelaktigt. Dekarboxyleringsreaktioner: Pyruvats omvandling till acetylCoA, isocitrats omvandling till -ketoglutarat, -ketoglutarats omvandling till succinylCoA eller oxaloacetats omvandling till fosfoenolpyruvat (i glukoneogenesen. 15. Cellerna är beroende av transportmolekyler för att alla metabola system ska fungera. a) Vilken transportör krävs för fettsyranedbrytningen, vad transporteras och vart? b) Vilken transportör krävs för fettsyrasyntesen, vad transporteras och vart? Svar: a) Karnitin transporterar fettsyror från intermembranutrymmet in till mitokondriematrix. b) AcetylCoA kan inte fritt lämna mitokondriematrix utan är beroende av reaktion med oxaloacetat (varvid citrat bildas) för transport till cytosolen. 16. a) Om du fick rådet att helt avstå från kolhydrater, hur skulle det då påverka din förmåga att bryta ner fett? Vad skulle bildas? b) Om du då fick rådet att se till att äta fettsyror med ett udda antal kolatomer, skulle det påverka din situation? Svar: a) Om för mycket fett bryts ner relativt kolhydrater kan ketonkroppar bildas. All acetylCoA kan ej brytas ner i citronsyracykeln (finns för få intermediärer att reagera med). b) Fettsyror med udda antal kolatomer bildar i sista steget propionylCoA som kan omvandlas till succinylCoA som är en intermediär i citronsyracykeln så det skulle leda till att mer oxaloacetat bildas som kan reagera med acetylCoA och ge ATP. 17. Beräkna hur mycket ATP som erhållits efter fullständig nedbrytning av laurat (dodekanoat, 12C). Svar: 78 ATP (6 acetylCoA = 6 varv i citronsyracykeln – för detta krävs 5 varv i -oxidationen) ATP från NADH: 6·3·2.5 + 5·1·2.5 = 57.5 ATP från FADH2: 6·1·1.5 + 5·1·1.5 = 16.5 GTP som kan omvandlas till ATP: 6 Kostnad att få in fettsyran i mitokondrien: -2 Totalt: 78 ATP REGLERINGEN: 18. Fosforylering/defosforylering är vanligt vid reglering av enzymaktivitet. a) Vad kallas ett enzym som fosforylerar ett annat protein? b) Vad kallas ett enzym som defosforylerar ett annat protein? c) Vilket ämne fungerar som fosfatdonator vid fosforyleringar? Svar: a) Kinas b) Fosfatas c) ATP 19. Nämn några olika sätt att reglera nyckelenzymers aktiviteter. Svar: *Allosterisk reglering – exempel ’feedbackinhibering’ *Olika isoformer i olika organ – Aktiviteten beror av organets behov * Reversibel kovalent modifiering – exempel fosforylering * Proteolytisk aktivering – exempel zymogenaktivering * Transkriptionell reglering – nysyntes av enzymer 20. Hur kan fosforylering/defosforylering vara så effektivt när det gäller att reglera ett enzyms aktivitet? Svar: *Två negativa laddningar introduceras i strukturen – ändrade elektrostatiska interaktioner – ändrad konformation – ändrad aktivitet. *Ändrar vätebindningsmönstret eftersom flera syren introduceras i strukturen– ändrad konformation – ändrad aktivitet. *ATP är fosfatdonator – energi tillförs vilket kan förändra jämviktsläget mellan proteinets olika konformationer. 21. Skissa schematiskt på signalvägen från aktivering av receptor till aktivering av effektorer (t ex ett enzym) och ge exempel på ’second messengers’. Svar: Frisättning av ’primary messenger’ – aktivering av receptor – frisättning av ’second messenger’ – aktivering av effektorer. ’Second messenger’ kan vara t ex cAMP, cGMP, Ca2+, inositol-1,4,5-trisfosfat (IP3) eller diacylglycerol (DG). 22. Vissa proteiner är inaktiva om inte en serin eller treonin fosforylerats. I vissa fall har dock dessa proteiner kunnat aktiveras genom att serinen eller treoninen genom lägesspecifik mutagenes bytts ut mot en glutamat. Förklara varför. Svar: Resultatet tyder på att det är en negativ laddning som krävs för att proteinet ska vara i sin aktiva konformation. 23. En muterad form av Gα-subenheten av ett G-protein gör GDP/GTP-utbyten även i frånvaro av en aktiverad receptor. Hur påverkar detta signaleringsvägen? Svar: Signaleringsvägen stimuleras om Gα-subenheten binder en GTP även i frånvaro av aktiverad receptor. 24. Vid muskelarbete utsöndras adrenalin för att stimulera nedbrytning av glukos för att ge muskeln energi. Glukos finns lagrat som glykogen. Adrenalin påverkar -adrenerga receptorn vilket leder till aktivering av Gα och cAMP. cAMP-fosfodiesteras är ett enzym som omvandlar cAMP till AMP. Hur påverkar inhibitorer av cAMP-fosfodiesteras glukosmobiliseringen i musklerna? Svar: Om cAMP-koncentrationen är fortsatt hög trots att receptorn inte längre aktiveras kommer PKA (proteinkinas A) också att fortsätta vara aktivt. Detta i sin tur leder till att enzymer som krävs för glukosmobiliseringen, glykogenfosforylas, också är aktiva och därmed får muskeln den energi som krävs men glykogendepåerna är ju begränsade och ny inlagring av glykogen kommer inte att ske.