Transkription 30 nov
advertisement

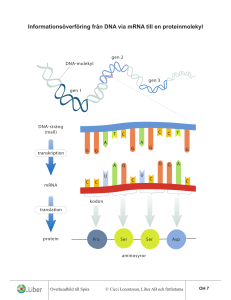
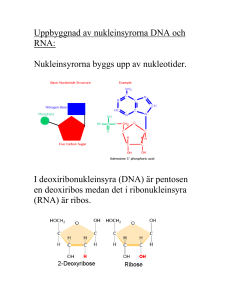
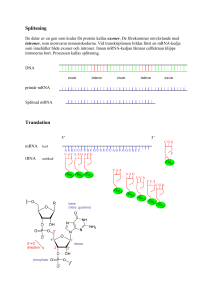
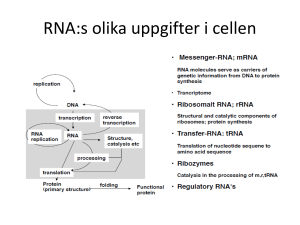
Molekylärbiologins centrala dogma m Replikation: Bassekvensen i DNA står för den genetiska informationen. När en cell ska delas måste DNA:t dupliceras – man måste få nytt DNA med exakt samma bassekvens som originalet. Vid proteinsyntes överförs informationen från DNA till aminosyrasekvens i två steg: Transkription: Bassekvensen i DNA översätts till bassekvens i mRNA. mRNA fungerar sedan som mall för proteinsyntesen. Translation: Bassekvens i mRNA översätts till aminosyrasekvens m h a tRNA. Processen sker på ribosomerna. En sekvens av tre baser utgör den GENETISKA KODEN och specificerar en viss aminosyra. Transkription = Översättning av bassekvensen i DNA till RNA OBS! Enklare process i prokaryota system - mycket mer komplicerat i eukaryota system! RNA: Tre huvudtyper: • tRNA • rRNA • mRNA transfer RNA ribosomalt RNA messenger RNA Alla tre är involverade vid proteinsyntesen: DNA mRNA Protein Transkription Translation rRNA tRNA I eukaryoter har man även funnit ytterligare en variant, snRNA (small nuclear): Involverade vid ’splicing’. Återkommer till detta! Funktioner hos RNA rRNA: Bygger tillsammans med proteiner upp RIBOSOMER. På ribosomerna sker proteinsyntesen. tRNA: Transporterar aminosyror från cytosolen till ribosomen. Ser till att rätt aminosyra hamnar på rätt plats i den växande polypeptidkedjan. mRNA: Utgör mall för proteinsyntesen. mRNA bildas genom avskrivning av DNA-sekvensen. Strukturen hos RNA - tRNA 3’ tRNA: Minsta typen av RNA, ca 80 nukleotider. Intramolekylära vätebindningar mellan kvävebaser gör att tRNA-molekylen blir klöverformad (2D). o Aminosyran som hämtas i cytosolen och transporteras till ribosomen binds till 3’-änden av tRNA-molekylen. o I antikodonloopen finns tre kvävebaser som basparar med mRNA och svarar för att aminosyrorna kommer i rätt ordning i proteinet. o Innehåller flera ovanliga, modifierade baser. Strukturen hos RNA - tRNA Tredimensionellt viker sig sedan tRNA så den blir L-formad. I den tredimensionella formen hamnar bindningsstället för aminosyran så långt som möjligt ifrån antikodonet. Olika tRNAmolekyler binder olika aminosyror – aminosyran måste ”stämma” med antikodonet. Strukturen hos RNA - tRNA Strukturen hos RNA - tRNA 5 3 Esterbindning En aminosyra kopplad på 3’-OH Strukturen hos RNA - rRNA Ribosom innehåller rRNA rRNA: Stora RNA molekyler som tillsammans med proteiner bygger upp ribosomer (2/3 av massan är RNA, 1/3 proteiner). 50S Ribosomerna består av en mindre och en större subenhet (30S och 50S). I eukaryoter (40S och 60S) 30S S = Sedimentationskonstanten = Svedberg 1 S motsvarar sedimentation på 10-13 s i en ultracentrifug (2 000 000 g) http://guweb2.gonzaga.edu/faculty/cronk/biochem/Rindex.cfm?definition=ribosome Strukturen hos RNA - mRNA mRNA mRNA: Innehåller bassekvensen som specificerar aminosyrasekvensen i ett protein. mRNA bildas när protein ska syntetiseras och bryts sedan snabbt ner för att nukleotiderna ska kunna återanvändas och för att cellen ska kunna kontrollera när ett visst protein syntetiseras. Bassekvensen i mRNA erhålls genom transkription (avskrivning) av bassekvensen i DNA. Strukturen hos RNA - mRNA Metylerad kvävebas Hos eukaryota organismer modifieras mRNA efter transkriptionen En ”cap” struktur bildas på 5’ änden och en längre sekvens med Adenin kvävebaser läggs till vid 3’ änden Ökar stabiliteten Roll mindre känt Ökar stabiliteten? Underlättar translationen? Strukturen hos RNA - mRNA mRNA innehåller informationen för ordningen på aminosyrorna i ett protein Tre nukleotider i följd kodar för en aminosyra Ex: UUU = Phe CAU = His AAG = Lys CGU = Arg CGC = Arg CGA = Arg CGG = Arg En aminosyra kan kodas av flera kodon = den genetiska koden är degenererad Strukturen hos RNA - mRNA mRNA innehåller start signal (AUG) för proteinsyntes Formylgrupp Strukturen hos RNA - mRNA mRNA innehåller stop signal för proteinsyntes Transkriptionen Transkriptionen katalyseras av RNA-polymeras, ett enzym som kopplar samman ribosnukleotider i riktning 5’ 3’. Hos E.coli består enzymet av fyra olika subenheter: α2ββ’σ ββ σ. σ-enhetens funktion är att hitta startstället, sedan dissocierar den och α2ββ’ katalyserar själva polymerisationen. Transkriptionen - mekanism Samma procedur som vid replikationen 1. Ny trifosfatnukleotid binder först till mallkedjan genom basparning i.e. vätebindning mellan två matchande kvävebaser 2. Hydroxylgruppen på C3 i den intilliggande nukleotiden gör en nukleofil attack på den innersta fosfor-atomen i den nya trifosfatnukleotiden. 3. PPi hydrolyseras och den nya nukleotiden sätts ihop kovalent med resten av RNA kedjan 4. Energikrävande process. Pyrofosfat (PPi) hydrolyseras varvid energi frigörs Transkriptionen - mekanism Nukleofil attack av OH på C3 5 3 3’,5’-fosfodiesterbindning 5 3 Ny nukleotid H-bindningar OBS! ribos i RNA deoxyribos i DNA Fosfaten är kopplad från C3 i kedjan till C5 i nästa socker, alltså 3’,5’-fosfodiesterbindning. Kedjan växer 5’ → 3’ – mRNA skrivs från 5’→ 3’ Transkriptionen RNA polymeras tar instruktionen från DNA Bassekvensen i mRNA är komplementär till den blåa DNA-strängen (templat kedjan) Den svarta DNA kedjan kallas för den kodande kedjan eftersom den har samma sekvens som mRNA förutom vid positionen för U i mRNA som är T i DNA kedjan Transkriptionen mRNA Transkriptionen - mekanism Mekanismen hos RNA polymeras liknar mekanismen för DNA-polymerasets: • Syntetiserar ny kedja 5’ 3’. • 3’-OH gör nukleofil attack på innersta fosfaten i trifosfatnukleotiden varvid pyrofosfat lämnar. • Tar instruktioner från en DNA-mall. Skillnader i funktion mellan DNA- resp. RNA-polymeras: • RNA-polymeraset behöver ingen primer. • RNA-polymeraset saknar exonukleas aktivitet. Gör inte lika mycket om det blir fel!!! Replikation – felet går vidare till nästa uppsättning celler Transkription – genererar endast ett tillfälligt fel i ett och annat protein Transkriptionen - start Transkriptionen startar vid s k promoterregioner. Dessa regioner är AT-rika: • • -35-regionen Pribnow-boxen (TATA-boxen) Ser lite olika ut vid olika promoter men båda sekvenserna måste finnas Transkriptionen - start RNA polymeraset måste vira upp DNA för att transkriptionen skall börja Ca 17 baspar Transkriptionen Transkriptionen sker i en transkriptionsbubbla som löper längst med DNA TRANSKRIPTIONSBUBBLA: DNA-helixen öppnas och stängs med samma hastighet så att det hela tiden är ca 17 baser oparade på den kodande DNA kedjan. Transkriptionen - terminering Transkriptionen fortgår till en termineringssignal nås. Terminering: Två sekvenser som innehåller många G och C viker RNA:t till en stabil hårnålsloop Därefter kommer en sekvens med flera U → destabilisering Destabilisering: A-U förbinds med 2 vätebindningar → RNA och DNA hålls inte ihop så bra längre – de separerar och RNA polymeraset släpper G och C basparar med 3 vätebindningar Transkriptionen - terminering Vissa gener kräver en annan faktor för att avsluta transkriptionen, ett protein som kallas rho. Rho känner igen ytterligare sekvenser i det nysyntetiserade transkriptet, binder till RNA:t vilket leder till att RNA-polymeraset lossnar. Processen kräver energi! Notera; I båda fallen ovan så är det signaler i det nysyntetiserade RNA:t som känns igen och inte signaler i DNA:t! Transkriptionen I prokaryoter translateras mRNA innan hela genen transkriberats I eukaryoter bildas ett primärt transkript som processas innan färdigt mRNA lämnar kärnan för transport till ribosomerna i cytosolen. Transkriptionen I prokaryoter syntetiseras RNA av ett polymeras men i eukaryoter finns det tre olika polymeraser Polymeraserna finns på olika ställen i cellkärnan och koder för olika RNA molekyler samt är olika känsliga för α-Amanitin som är ett gift från svampen Amanita phalloides Lömsk flugsvamp Transkriptionen Hos eukaryoter RNA polymeras I katalyserar formation av rRNA som skall ingå i ribosomen Från transkriptionen fås pre-rRNA som innehåller tre olika rRNA molekyler (18S, 5.8S samt 28S) Modifiering av både ribos- och kvävebasenheter (röda streck). Ex metylering Klyvning → Färdigprocessat rRNA 40S 60S En annan rRNA-molekyl (5S) som skall ingå i 60S transkriberas av RNA polymeras III Transkriptionen Hos eukaryoter RNA polymeras III katalyserar formation av tRNA Processning av tidigt tRNA transkript Borttag av nukleotider vid 5’ och 3’ ändan Tillägg av CCA vid 3’ änden Borttag av en 14 nukleotid lång intron → splicing Modifiering av socker- och kvävebasenheter (viktigt för funktionen) Transkriptionen Hos eukaryoter RNA polymeras II katalyserar formation av mRNA För att RNA-polymeraset ska hitta rätt startställe på DNA templatet krävs ett antal transkriptionsfaktorer; TFII A, B, D, E, F, H. (TF = transkriptionsfaktorer, II – RNA polymeras II) TFIID binder till TATA-boxen via en av sina subenheter, TBP (’TATA-box binding protein’). Detta leder till konformationsförändringar i DNA TFIIA och TFIIB binder in till TBP:s yta. Transkriptionen Efter att TFIIB bundit in byggs komplexet på med TFIIF, E och H och RNA polymeras II TFIIH öppnar DNA helixen och fosforylerar den Cterminala domänen (CTD) av polymeraset → polymeraset lämnar promotorn och börjar transkriptionen Transkriptionen Hos eukaryoter RNA polymeras II katalyserar formation av mRNA 5’ änden stabiliseras av Cap För att avsluta transkriptionen känner ett specifikt endonukleas igen sekvensen AAUAAA och klyver RNA nedströms En svans AAAAA(A)n kopplas på av poly(A)polymeras Splicing Färdigt mRNA Transkriptionen – splicing av mRNA • De primära mRNA transkripten i eukaryoter består av EXONER och INTRONER. • Exonerna innehåller den genetiska informationen som kodar för protein - 2% av vårt genmaterial • Intronerna (- 98 % av vårt genmaterial) används inte som mall för proteinsyntes utan dessa klipps bort från mRNA – processen kallas för splicing • I medeltal innehåller en gen hos människan 8 introner men det kan finnas upp till 100 stycken • Ofta kodar exonerna för olika funktionella domäner eller subenheter i ett protein • Genom olika (alternativ) splicing kan olika varianter av mRNA bildas från en och samma kodande region på DNA Transkriptionen – splicing av mRNA Processen då introner avlägsnas och exoner sammanfogas kallas ’splicing’. Exon Exon Splicing mRNA före splicing mRNA efter splicing Transkriptionen – splicing av mRNA Alternativ splicing genererar mRNA som är templat för olika proteinformer Splicing 1 av mRNA genererar ett membranbundet protein Splicing 2 av mRNA genererar ett protein som saknar den membranbindande delen Splicing Mekanism: En intron startar alltid med GU och slutar med AG. Den har också alltid ett internt förgreningssäte med ett A. Exon 1 Exon 2 Splicing Stegvis: 1. 2’-hydroxylgruppen på A i förgreningssätet gör en nukleofil attack på fosforn i fosfodiesterbindningen som förbinder exon 1 och intronen. Bindningen bryts. 2. 3’-hydroxylgruppen på exon 1 attackerar fosfodiesterbindningen mellan exon 2 och intronen varvid en ny fosfodiesterbindning bildas mellan exon 1 och exon 2. Splicing Katalyseras av ett komplex, SPLICEOSOMEN, som består av proteiner och snRNA (sn = small nuclear). U 1, 2, 4, 5, 6 är snRNA som har en viktig funktion vid splicing: U1 binder till 5’-sätet och U2 till förgreningssätet Ett komplex av U4-U5-U6 binder in och SPLICEOSOMEN är komplett U6 binder in till 5’-sätet och knuffar bort U1 och U6-U2 interagerar med varandra och knuffar bort U4. 2’-hydroxylgruppen på A i förgreningssätet attackerar 5’-sätet varpå bindningen bryts. U5 håller de två exonerna nära varandra hydroxylgruppen i 5’-sätet attackerar 3’-sätet → färdigsplicad exon Splicing Alternativ splicing: Viktigt för ett begränsat genmaterial kan ge upphov till många olika proteiner. Typexempel: Antikroppsproduktion Bananflugor har ca 14 000 gener och en människa har ungefär dubbelt så många ca 24 000 Bananflugans huvudsysselsättning är att finna en banan och vi har många fler funktioner. Tittar man på antalet mRNA vi bildar är det ca 150 000 alltså 6 ggr mer än generna. Slutsats: Genmaterialet kan användas för att ge flera olika mRNA transkript Notera; En stor andel av alla genetiska sjukdomar (speciellt neurologiska) orsakas av mutationer som påverkar var gener ’splicas’. Man tror att över 70% av alla gener är föremål för alternativ splicing för att producera funktionellt olika proteiner. Grundtesen, en gen = ett protein stämmer alltså inte. Vårt genmaterial skiljer sig motsvarande 1-2% från chimpanserna. Om vi inte hade alternativ splicing så skulle inte skillnaderna i proteinuttryck vara så stor som den är mellan människa och chimpans. Ange mRNA-sekvensen som motsvarar följande DNA sekvens 5’ CGA TGT TAG CGT-3’ Templat kedjan Ange mRNA-sekvensen som motsvarar följande DNA sekvens 5’ CGA TGT TAG CGT-3’ Templat kedjan 3’ GCU ACA AUC GCA-5’ mRNA från 3’-5’ SVAR: 5’ ACG CUA ACA UCG 3’