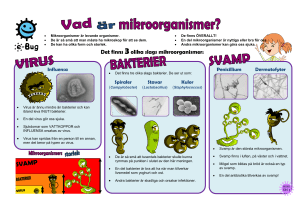
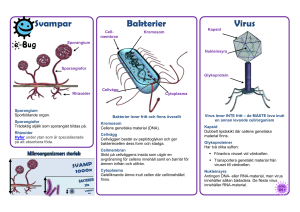
Mikrobiologisk barriär- verkan vid konstgjord grundvattenbildning
advertisement

VA-Forsk rapport Nr 2006-10 Mikrobiologisk barriär­ verkan vid konstgjord grundvattenbildning – en litteraturstudie om påverkande faktorer Kajsa Engblom Måns Lundh VA-Forsk VA-Forsk VA-Forsk är kommunernas eget FoU-program om kommunal VA-teknik. Programmet finansieras i sin helhet av kommunerna, vilket är unikt på så sätt att statliga medel tidigare alltid använts för denna typ av verksamhet. FoU-avgiften är för närvarande 1,05 kronor per kommuninnevånare och år. Avgiften är obligatorisk. Nästan alla kommuner är med i programmet, vilket innebär att budgeten årligen omfattar drygt åtta miljoner kronor. VA-Forsk initierades gemensamt av Svenska Kommunförbundet och Svenskt Vatten. Verksamheten påbörjades år 1990. Programmet lägger tonvikten på tillämpad forskning och utveckling inom det kommunala VA-området. Projekt bedrivs inom hela det VA-tekniska fältet under huvudrubrikerna: Dricksvatten Ledningsnät Avloppsvattenrening Ekonomi och organisation Utbildning och information VA-Forsk styrs av en kommitté, som utses av styrelsen för Svenskt Vatten AB. För närvarande har kommittén följande sammansättning: Anders Lago, ordförande Olof Bergstedt Roger Bergström Daniel Hellström Stefan Marklund Mikael Medelberg Anders Moritz Peter Stahre Jan Söderström Göran Tägtström Agneta Åkerberg Södertälje Göteborgs VA-verk Svenskt Vatten AB Stockholm Vatten AB Luleå Roslagsvatten AB Linköping VA-verket Malmö Sv Kommunförbundet Borlänge Falkenberg Steinar Nybruket, adjungerad Thomas Hellström, sekreterare NORVAR, Norge Svenskt Vatten AB Författarna är ensamma ansvariga för rapportens innehåll, varför detta ej kan åberopas såsom representerande Svenskt Vattens ståndpunkt. VA-Forsk Svenskt Vatten AB Box 47607 117 94 Stockholm Tfn 08-506 002 00 Fax 08-506 002 10 [email protected] www.svensktvatten.se Svenskt Vatten AB är servicebolag till föreningen Svenskt Vatten. VA-Forsk Bibliografiska uppgifter för nr Rapportens titel: Mikrobiologisk barriärverkan vid konstgjord grundvattenbildning – en litteraturstudie om påverkande faktorer Title of the report: Microbiological barrier in artificial recharge of ground water – a review Rapportens beteckning Nr i VA-Forsk-serien: 2006-10 Författare: Kajsa Engblom, Måns Lundh, Göteborgsregionens kommunalförbund (GR) VA-Forsk-projektnr: 24-103 Projektets namn: Mikrobiologisk barriärverkan i konstgjord grundvattenbildning – litteraturstudie Projektets finansiering: VA-Forsk Rapportens omfattning Sidantal: Format: 51 A4 2006-10 Sökord: Dricksvatten, mikrobiologisk barriär, konstgjord grundvattenbildning, redukt­ ion, patogen, bakterier, virus Keywords: Drinking water, microbiological barrier, artificial recharge, groundwater, reduction, pathogen, bacteria, virus Sammandrag: Litteraturstudien redovisar vilka faktorer som påverkar den mikrobiologiska barriären vid konstgjord grundvattenbildning och hur de påverkar. Studien sammanfattar reduktionen av patogener vid olika fullskaleförsök. Abstract: The review accounts for the factors influencing the microbiological barrier at artificial recharge of groundwater. A summery of the reduction of pathogens for different full scale studies is made. Målgrupper: VA-ansvariga, tillsynsmyndigheter, VA-konsulter, driftpersonal på vattenverk med konstgjord grundvattenbildning Omslagsbild: Vy över Dösebacka vattenverk, Kungälv. Fotograf: Kajsa Engblom Rapporten beställs från: Finns att hämta hem som pdf-fil från Svenskt Vattens hemsida www.svensktvatten.se Utgivningsår: 2006 Utgivare: Svenskt Vatten AB © Svenskt Vatten AB Grafisk formgivning: Victoria Björk, Svenskt Vatten Förord Litteraturstudien genomfördes mellan april till november 2005 med finansiering från VA-Forsk. Rapporten riktar sig till alla intresserade av forskning och utveckling av konstgjord grundvattenbildning eller utformning och drift av en anläggning med infiltration som ett reningssteg. Studien ingick i ett större projekt om mikrobiologisk barriärverkan vid konstgjord grundvattenbildning som pågått i Göteborg 2003-2005. Projektet syftade till att undersöka tillämpligheten av konstgjord grundvattenbildning i Gråbodeltat vid sjön Mjörn. Vi tackar alla som bidragit till produktionen av denna rapport och genomförandet av studien. Ett speciellt tack till Olof Bergstedt, Göteborgs va-verk, för värdefulla kommentarer och Emma Holmström, GR, för synpunkter på utformningen av rapporten. Kajsa Engblom Måns Lundh Göteborg 2006-01-31 Göteborg 2006-01-31 Innehåll Förord. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 Sammanfattning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7 Summary. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8 1 Introduktion. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9 1.1 Bakgrund . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9 1.2 Syfte. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10 1.3 Metod. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10 1.4 Begränsningar . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10 1.5 Rapportens disposition. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10 2 Konstgjord grundvattenbildning – metodbeskrivning. . . . . . . . . 11 2.1 Begrepp. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11 2.2 Anläggningstyper . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11 2.3 Reningsprocesser i marken vid konstgjord grundvattenbildning – omvandling från ytvatten till grundvatten. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12 2.4 Igensättningar. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13 2.5 Fördelar och nackdelar med konstgjord grundvattenbildning. . . 13 3 Patogener och indikatororganismer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13 3.1 Patogena organismer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13 3.1.1 Virus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14 3.1.2 Bakterier. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14 3.1.3 Protozoer. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15 3.2 Vattenburna patogener och lagstiftning. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15 3.3 Indikatororganismer. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17 3.3.1 Bakteriofager som indikatororganismer och modellorganismer . . . . 18 4Studier om mikrobio­logisk barriärverkan vid konstgjord grundvatten­bildning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19 5 Påverkande faktorer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20 5.1 Översikt. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20 5.2 Avskiljning. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21 5.3 Inaktivering och avdödning. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22 5.4 Relationen mellan inaktivering och avskiljning. . . . . . . . . . . . . . . . . . . . . . . . 22 6De olika faktorernas inverkan på reduktionen av patogener . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23 6.1 Egenskaper hos mikroorganismen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23 6.1.1 6.1.2 6.1.3 6.1.4 6.1.5 Koncentration. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23 Organismtyp . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23 Storlek och form. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23 Laddning hos mikroorganismerna. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24 Hydrofobicitet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24 6.2 6.2.1 6.2.2 6.2.3 6.2.4 6.3 6.3.1 6.3.2 6.3.3 6.3.4 6.4 6.4.1 6.4.2 6.4.3 6.4.4 6.4.5 Egenskaper hos vattnet. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24 pH . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24 Temperatur . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25 Innehåll av organiskt material . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26 Jonkoncentration och jonsammansättning i vattnet. . . . . . . . . . . . . . . . . . . . 27 Egenskaper hos marken och infiltrationsmaterialet. . . . . . . . . . . . . . . . . 28 Partikel och porstorlek . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28 Mineral- och Jordsammansättning. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28 Laddning hos marken. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29 Hydrofobicitet. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29 Anläggningens utformning och drift . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30 Uppehållstid. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30 Transportsträcka mellan infiltration och uttag. . . . . . . . . . . . . . . . . . . . . . . . . . . . 31 Grundvattenhastighet. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32 Vattenmättnadsgrad. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32 Biohud och naturliga mikroorganismer. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33 7 Kvantifiering av de dimensionerande faktorerna . . . . . . . . . . . . . . . . 33 7.1 Inledning. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33 7.2 Metod. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34 7.3 Reduktion i förhållande till transportsträckan. . . . . . . . . . . . . . . . . . . . . . . . . 34 7.4 Reduktion i förhållande till uppehållstid. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34 7.5 Reduktionsfaktor . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36 7.6 Modellorganismer. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37 7.7 Diskussion av kvantifierings­studien och jämförelse med lagstiftning internationellt. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37 7.8 Begränsningar . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38 8 Sammanfattning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39 8.1 Påverkande faktorer. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39 8.2 Kvantifieringsstudien. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40 9 Forsknings- och utvecklingsbehov. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41 Referenser. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43 Bilaga A: Sökord. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48 Bilaga B: Data från fältstudier . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49 Sammanfattning Den mikrobiologiska säkerheten mot vattenburna sjukdomsutbrott är en viktig funktion vid produktion av dricksvatten för att förhindra sjukdomsfall. En ökad kännedom om mikrobiologiska föroreningskällor, ett vakuum kring lagstiftning med krav på råvattenkvalitet samt brist på kunskap om virus motiverar forskning och studier om mikrobiologiska säkerhetsbarriärer vid dricksvattenberedning. Konstgjord grundvattenbildning är en förstärkning av den naturliga tillgången av grundvatten genom infiltration av ytvatten. Det finns flera olika metoder som används i Sverige, varav bassänginfiltration är den vanligaste. Reningen av vattnet sker i den bildade biohuden i botten av bassängerna, samt i jordlagren i marken, som fungerar som mikrobiologisk barriär. Litteraturstudien granskar de senaste rönen om mikrobiologisk barriär i konst­ gjord grundvattenbildning och mikroorganismers transport i mark och grundvatten för att identifiera vilka faktorer som är av betydelse för reduktionen och hur de påverkar. Det är egenskaper hos mikroorganismen, vattnet och marken som avgör hur och till vilken grad reduktionen sker. Även hur infiltrationsanläggningen är utformad och drivs påverkar. I studien har ett försök gjorts till att kvantifiera den mikrobiologiska barriärverkan vid olika uppehållstider och avstånd från infiltrationspunkten, utifrån resultat från undersökningar som gjorts i fullskaliga anläggningar och försöksområden i olika länder. Det är stor skillnad på reduktionen beroende på akvifärens egenskaper, då främst avseende grundvattenhastigheten och hur anläggningen drivs, och noggranna lokala undersökningar måste utföras. Enligt följande tabell som sammanställts med hjälp av fullskaleförsöken ger olika uppehållstider och avstånd mellan infiltrations- och uttagspunkt varierande effektivitet på barriären: Långsam akvifär (1–3 m/dag) Uppehållstid (dygn) Snabb akvifär (>25 m/dag) Avstånd (m) Uppehållstid (dygn) Avstånd (m) Erhållen Reduktion (log10 ) >5 10 ca 1 25 3 >14 15 ca 1,5 40 4 >25 20 ca 2 80 5 >35 30 ca 3 >80 6 Efter 14 dagar, vilket är gränsen för att klassificera ytvatten som grundvatten, är reduktionen mellan 4-8 log10, beroende på akvifärens förutsättningar. Eftersom avståndet mellan infiltrationspunkten och uttaget är en lika relevant parameter, enligt denna studie, skulle Livsmedelsverkets rekommendation om 14 dagar kunna kompletteras med ett rekommenderat avstånd. Denna rekommendation skulle kunna hamna på ca 40 m, vilket är tillräckligt för en reduktion på mellan 4-8 log10 av virus. Summary The microbiological barrier for protection against pathogens (microorganisms causing disease) is an important function in the production of drinking water. The increasing awareness of potential causes for microbiological contamination, a vacuum in Swedish regulation on quality of the raw water, as well as a lack of knowledge about virus motivates research and studies about microbiological safety barriers in drinking water production. The review gives a short introduction of artificial recharge and the different methods used in Sweden. The processes that take place in the different soil layers acts as the microbiological barrier in artificial recharge. One difficulty with drinking water production through artificial recharge is quantifying the reduction of micro­ organisms and other substances. The review examines the latest results and discussions about the microbiological barrier and microorganisms transport in soil and groundwater in order to identify the factors affecting the reduction and how. There are several factors, including characteristics of the microorganism, the water and the soil influencing the reduction. The design and management of the plant are also of significance. An attempt of quantifying the microbiological barrier at different travel times and distances from the injection/infiltration point has been made to facilitate the assessment and construction of a plant for artificial recharge. The results from full scale studies, compiled in the following table, show that there is a great variety in the reduction of microorganisms depending on the characteristics of the aquifer and especially the velocity of the groundwater influence the reduction. This emphasizes the importance of careful and accurate local studies. Slow aquifer (1–3 m/day) Travel time (days) Rapid aquifer (>25 m/day) Distance (m) Travel time (days) Distance (m) Barrier removal efficiency (log10 ) >5 10 ca 1 25 3 >14 15 ca 1,5 40 4 >25 20 ca 2 80 5 >35 30 ca 3 >80 6 After 14 days, which is the limit for defining infiltrated water as groundwater in Sweden, the reduction of virus is between 4–8 log10 according to this review. Since the distance between the infiltration and abstraction point is an equally relevant parameter, a recommendation of a minimum distance could complement the National Food Administration’s recommendation of 14 days. This recommendat­ ion could be a distance of approximately 40 m, which is sufficient for a reduction of 4–8 log10. av naturliga partiklar i bakteriestorlek och en logavskiljning på 4,7–5,5 för tillsatta bakteriofager (Persson et al. 2005). Klorering ska ge avdödning av 99 % av bakterierna, men det är diskuterat om klorering verk­ ligen ger den effektiviteten på mer tåliga virus och parasiter. Livsmedelsverket (2004) innefattar långsam­ filtrering som en mikrobiologisk barriär men nämner inga kvantitativa värden på vad reduktionen är. Det problematiska med dricksvattenförsörjning genom konstgjord grundvattenbildning är svårigheten att generellt kvantifiera reningseffektiviteten, dvs. re­ duktionen, av olika ämnen och mikroorganismer (den mikrobiologiska barriärverkan). Barriärverkan i konstgjord grundvattenbildning beror mycket på förhållanden som varierar mellan olika anläggningar och kan inte bestämmas lika enkelt som för ytvatten­ verk. Fysikaliska och geologiska förutsättningar, såsom infiltrationsvattnets och markens egenskaper samt processutformning skiljer sig mellan olika anläggningar. De fysikaliska, kemiska och biologiska processer för rening, och de faktorer som styr dessa inom konst­ gjord grundvattenbildning, är inte väl undersökta i Sverige. Därför krävs en djupare kunskap om överlevnad och avskiljning av patogener i mark och vatten, för att utveckla effektiva bestämmelser och föreskrifter för att förhindra förorening av dricksvatten. Det krävs även en utveckling av nya metoder för analys och övervakning för en säker behandling (Jörgensen et al. 2001). Fryklund m.fl. (1994) menar på att den forskning som bedrivits inom området utomlands och de resultat som erhållits är inte direkt tillämpbara på svenska förhållanden. De mikroorgan­ ismer och det organiska materialet som ska avskiljas är av olika typ, de hydrogeologiska förhållandena skiljer sig åt osv. och motiverar att svenska undersökningar behövs. Det pågår ett forskningsprojekt i Göteborg om mikrobiologisk barriärverkan i omättad zon vid konst­ gjord grundvattenbildning. Resultaten kommer att presenteras som en VA-Forsk rapport. Denna litter­ aturstudie är utförd som bakgrund och kunskapsinhämtning till projektet för att ge förståelse för pro­ blematiken, samt uppslag på vad framtida svensk forskning inom området bör inriktas på. 1 Introduktion 1.1 Bakgrund Vattenburna sjukdomsutbrott förekommer i Sverige liksom i resten av världen. Under åren 1980–2004 rapporterades 142 vattenburna utbrott med totalt 63 000 sjukdomsfall runtom i landet. I genomsnitt rapporteras nästan ett vattenburet utbrott med minst 1000 drabbade per år. Campylobakter har varit det smittämne man funnit flest gånger, följt av norovirus och Giardia Lamblia, men i merparten av de vatten­ burna utbrotten har smittan inte lyckats identifieras (Smittskyddsinstitutet 2005). Den mikrobiologiska säkerheten mot patogener (sjukdomsframkallande mikroorganismer) som bakterier, virus och protozoer av fekalt ursprung är därmed en viktig funktion vid produktion av dricksvatten. En ökad kännedom om mikrobiologiska föroreningskällor, ett vakuum kring lagstiftning med krav på råvattenkvalitet samt bristen på kunskap om virus motiverar forskning och studier om mikrobiologiska säkerhetsbarriärer vid dricksvattenberedning. De svenska föreskrifterna för dricksvattenproduktion innehåller i dag inga specifika och kvant­ itativa krav på hur effektiva de mikrobiologiska säkerhetsbarriärerna ska vara i att avskilja sjukdoms­ framkallande mikroorganismer. Enligt vägledningen till föreskrifterna ses en låg turbiditet (<0,1 FNU) och en 99 % (2 log10) inaktivering av bakterier som min­ iminivå för att beredningssteget ska räknas som en barriär (Livsmedelsverket 2004). För kemisk fällning och snabbfiltrering ligger avskiljningen av virus och fekala indikatorbakterier, som tillsätts i höga koncentrationer, på mellan 3 och 4 log10, men ner till 1–2 log10 för naturliga partiklar i bakterie eller protozo­ storlek. Ultrafiltrering, med membran på 100 nm, visade sig i försök ge en avskiljning på 98–99,9 % Se kapitel 3 för vidare förklaring av begreppet patogen. Avföring, exkrementer. Med en mikrobiologisk säkerhetsbarriär menas en anordning eller åtgärd i vattenverket som motverkar förekomst av sjukdomsframkallande virus, bakterier och parasitära protozoer i dricksvattnet (Livsmedelsverket 2004). Reduktion av naturligt organiskt material och mikroorgan­ ismer i konstgjord grundvattenbildning. Måns Lundh, Emma Holmström, Jonas Långmark, Henrik Rydberg, under arbete 2005. och inte vidare till de källor som litteraturstudien i sin tur hänvisar till (t.ex. Stevik et al. 2004; Leclerc et al. 2000 och Schjiven 2001). I vissa fall har inte andrahandskällor undersökts (detta angivs i texten som t.ex. Nasser et al. 1993, citerad i Pang et al. 2003) p.g.a. tidsbrist. 1.2 Syfte Syftet med litteraturstudien är att granska de senaste rönen om mikrobiologisk barriär i konstgjord grund­ vattenbildning och mikroorganismers transport i marken för att identifiera vilka faktorer och mätbara parametrar som är av betydelse för reduktionen och hur de påverkar. Studien ska dessutom göra en ansats för att klargöra förhållanden mellan de parametrar som påverkar reduktionen av sjukdomsframkallande mikroorganismer och skillnader i reduktionen mellan virus, bakterier och protozoer. Studien ska förhoppningsvis kunna förenkla den komplicerade process som en bedömning och dimensionering av en anläggning för konstgjord grund­ vattenbildning är med avseende på den mikrobiolog­ iska säkerheten, genom att kvantifiera den mikrobiologiska barriärverkan vid olika uppehållstider och transportsträckor. 1.4 Begränsningar En allmän begränsning är urvalet av artiklar som bygger på en subjektiv bedömning av de som funnits i databaserna. Alla artiklar inom området har inte kunnat granskas och analyseras, det har tiden för projektet inte medgivit. Förhållandena skiljer sig i Sverige mot förhållandena i studerad litteratur med avseende på vattenkvaliteten och markförhållanden, vilket medför att jämförelsen av resultat från studier med praktiska erfarenheter från anläggningar begränsas. De faktorer som studerats mest noggrant är de som gått att undersöka i pilotförsök och i labbskala vilket gör att flera faktorer, bl.a. omättad zons be­ tydelse, som kan vara avgörande på en fullskalenivå, är dåligt undersökta. Applicering av resultat och slut­ satser från försök i pilotskala på verkligheten är behäftat med osäkerheter och ett samspel mellan olika faktorer kan medföra att förhållanden blir annorlunda. Begränsningar med metoden och genomförandet av kvantifieringen diskuteras i kapitel 10. 1.3 Metod Arbetet har utförts som en litteraturstudie av veten­ skapliga tidskrifter- papers, rapporter och handböcker. Två huvudsakliga typer av litteratur har använts för beskrivningen och kvantifieringen av hur de olika faktorerna påverkar reduktionen; vetenskapliga art­ iklar som redovisar utförda försök och resultat samt litteraturstudier liknande denna som diskuterar den mikrobiologiska barriärverkan utifrån tidigare forskning. Metoden för kvantifieringen är beskriven i kapitel 10. Artiklarna har främst fåtts från olika elektroniska databaser (Elsevier- ScienceDirect, ProQuest, Springer Link m.fl.) med tidskrifter i fulltext, samt med referenslitteratur, handböcker m.m. Arbetet är med andra ord i huvudsak en elektronisk litteraturstudie där äldre referenser inte har prioriterats p.g.a. svårig­ heten att få tag på dem, utan de flesta av referenserna är från det senaste decenniet och finns publicerade elektroniskt. De sökord som använts för att söka relevant material finns i Bilaga A och för varje artikel som använts har en noggrann genomgång av referens­ listan gjorts för att finna ytterligare material. För den information som hämtats från andra litter­ aturstudier har referens endast gjorts till aktuell studie 1.5 Rapportens disposition Rapportens huvudkapitel om de påverkande faktor­ erna (kapitel 5 och 6) föregås av en kort introduktion till konstgjord grundvattenbildning och sjukdoms­ alstrande mikroorganismer, (kapitel 2 och 3) samt en beskrivning av hur studier utförs på området (kapitel 4). Kvantifiering av den mikrobiologiska barriären behandlas i ett separat kapitel (7) och rapporten avslutas med en sammanfattning (kapitel 8) och en diskussion om vidare forskningsbehov (kapitel 9). 10 kylvatten, grundvattennivåkontroll och som hydraul­ isk spärr (Hanson 2000). 2 Konstgjord grundvattenbildning – metodbeskrivning 2.2 Anläggningstyper Kapitlet ger endast en översikt av processen för konst­ gjord grundvattenbildning. För mer ingående beskrivningar hänvisas till litteratur i referenslistan; Bouwer (2002) och speciellt VA-Forsk rapporterna Hanson (2000), Fryklund m.fl. (1994) samt Sundlöf och Kronqvist (1992). Det tre infiltrationsmetoder som används i Sverige är: • Bassänginfiltration – Avledning eller pumpning av vatten från sjöar eller vattendrag till bassänger på sand- och grusavlagringar, där det får infiltrera. Bassängerna innehåller vanligtvis filtersand ovanpå de naturliga jordlagren och på ytan bildas ofta ett biologiskt aktivt skikt, en s.k. biohud som medverkar vid reningen. Det infiltrerade vattnet pumpas sedan upp som grundvatten. Detta är den teknik som är vanligast i Sverige (se fig. 2-1). • Inducerad infiltration – Pumpning ur grundvatten­ brunnar som står i hydraulisk förbindelse med en sjö eller vattendrag. Uttaget av vatten gör att ytvattnet infiltreras genom bottensedimenten eller strandkanten. Reningsprocesserna sker mer okont­ rollerat än vid bassänginfiltration. • Djupinfiltration – Om täta lerlager förhindrar metoderna ovan kan infiltration av ytvatten ske direkt i brunnar. Metoden kräver normalt för­ behandling, för att kompensera att ingen rening sker i den omättade zonen samt för att förhindra igensättningar av brunnarna (Hanson 2000). 2.1 Begrepp I Sverige är vattenförsörjningen till ca 50 % baserad på ytvatten och resterande 50 % utgörs av lika delar naturligt och konstgjort grundvatten. För att förstå konceptet med konstgjord grundvattenbildning kan följande begrepp definieras: • Grundvattenbildning: ”den naturliga eller artificiella process, där vatten tillförs utifrån (nederbörd, ytvatten, bevattning etc.) till den mättade zonen av en akvifär antingen direkt till formationen eller indirekt via en annan formation” (Hanson 2000). • Akvifer/Akvifär: ”en geologisk formation som kan avge vatten i betydande mängd” (Hanson 2000). • Konstgjord grundvattenbildning innebär således den artificiella process som nämns i definitionen ovan. Akvifären fylls på genom att ytvatten läggs i bassänger eller diken alternativt sprinklas över marken för att det ska infiltrera ner till de naturliga grundvattenmagasinen. Det finns ytterligare en metod av konstgjord grundvattenbildning, men den används inte i Sverige idag. Det pågår däremot ett utvecklingsprojekt sedan 2003 i Katrineholm för att undersöka metoden och möjligtvis använda den för framtida vattenförsörjning av Katrineholms kommun (Djurberg 2005). • Sprinklerinfiltration – Metoden liknar bassänginfiltration men vattentillförseln till akvifären sker genom att sprida ytvatten i stora mängder genom perforerade ledningar direkt på marken (Hanson 2000). En fördel med denna metod är att ingreppet i naturen blir minimalt. Konstgjord grundvattenbildning används för flera olika syften. Det vanligaste är för dricksvattenförsörjning, vilken är målet som denna studie fokuserar på. Inom dricksvattenförsörjning kan konstgjord grundvattenbildning användas med syftet att rena ytvatten genom infiltration, men även med syftet att bidra med förstärkning till den naturliga grundvattenbildningen och vattenbehandling genom bland­ ning av yt- och grundvatten. Men även andra användningsområden förutom dricksvattenförsörjning förekommer; lagring av vatten kortare eller längre tid, grundvattenvärme, temperaturutjämning för En typisk svensk ”median” anläggning beskrivs i Sundlöf och Kronqvist (1992). Bassänginfiltration, som är den vanligaste metoden, sker med ytvatten från en sjö med någon typ av förbehandling, t.ex. mikrosilning. Vattnet infiltreras med en hastighet av 2 m/dygn, uppehållstiden är 1–2 månader och 11 Ytvatten Uttagsbrunn Infiltrationsbassäng Omättad zon Grundvattenyta Mättad zon Figur 2‑1. Schematisk beskrivning av bassänginfiltration. avståndet till brunnarna från bassängerna ca 300 m. Normalt rensas bassängerna från en alltför kraftig biohud ca 1–2 ggr/år. Kronqvist 1992). Naturliga reningsprocesser som sker i marken skiljer sig i de olika markskikten. I de översta lagren, förnan och humuslagren, avskiljs organiskt material genom hydrofob adsorption på sandkornen och vattnet är surt (pH 4–5). I blekjordslagret, djupare ned, löser det sura vattnet ut mineraler och vattnets pH, alkalinitet och hårdhet ökar. I det djupare lagret, rostjordslagret, avskiljs organiskt material genom elektrostatisk adsorption och utflockning med pos­ itivt laddade järn- och aluminium(hydr)oxider och vattnet blir klarare. Vattnets pH, alkalinitet och hård­ het ökar genom jonbytesprocesser ännu längre ned i marken, den s.k. C-horisonten, och nere vid grund­ vattenzonen liknar vattnet alltmer grundvatten i sin karaktär (Hanson 2000). Detta är vad som sker i naturliga marklager med infiltration av opåverkat regnvatten. Vad som sker vid konstgjord infiltration med väsentliga skillnader i mark- och vattenkaraktär samt en högre hydraulisk belastning är inte helt klar­ lagt. Reningsprocesserna skiljer sig till viss del mellan de olika metoderna för konstgjord grundvattenbildning nämnda ovan, men grundprinciperna är desamma som för naturlig grundvattenbildning. I de översta lagren vid bassänginfiltration sker en fastläggning av suspenderat material, avskiljning av organiskt material och minskning av bakterier och virus. Djupare ner i filterbädden sker ytterligare avskiljning av organiskt material. Under bassängen i de naturliga jordlagren tillförs vattnet joner, bl.a. 2.3 Reningsprocesser i marken vid konstgjord grundvattenbildning – omvandling från ytvatten till grundvatten Ett ytvatten är mycket varierande i sin karaktär, både mellan olika sjö- och vattendragsystem och mellan årstiderna. Innehållet av organiskt material, syre, salter och mineraler, samt levande organismer kan skilja sig mycket från grundvatten och från dricksvatten. Ett grundvatten karakteriseras av låga halter organiskt material och partiklar, låg syrehalt och förekomst av järn, mangan och utlösta salter. Antalet mikroorganismer är mycket reducerat. (Sundlöf & Kronqvist 1992). Vid produktion av dricksvatten är det önskvärt med en komposition av grundvattenkaraktär. Marken fungerar som ett medium för att rena vatten från olika naturligt förekommande ämnen och föroreningar och ombildar ytvatten till grundvatten. Det sker genom ett flertal processer som mekanisk filtrering, kemiska reaktioner i syrerika och syre­ fattiga miljöer och biologisk nedbrytning (Sundlöf & 12 Ca2+, Mg2+, och K+, i jonbytesprocesser. Vattnets pHvärde, alkalinitet och hårdhet ökar och det organiska materialet minskar. När vattnet når grundvattnet temperaturutjämnas det och vid syrebrist kan järn och manganhalten öka. För inducerad infiltration avskiljs en del av det organiska materialet i strandoch bottensedimenten och i finsedimenten beroende på tillgången på syre. Den syrefria miljön som ofta uppstår närmare grundvattnet bidrar till att järn och mangan löser ut och vattnet måste ofta efterbehandlas med avseende på detta (Hanson 2000). Det är viktigt att uppehållstiden mellan infiltrat­ ionen och uttaget är väl avvägd. Tillräckligt lång tid krävs för att avskiljning av organiskt material och reduktionen av bakterier och virus ska bli fullgod. Men en för lång uppehållstid kan leda till att syrebrist uppstår och att ämnen som järn och mangan kan lösas ut och vattnet kräver efterbehandling. 2.5 Fördelar och nackdelar med konstgjord grundvattenbildning De fördelar som finns med dricksvattenproduktion genom konstgjord infiltration är • Förenklad reningsprocess • Stor magasinsvolym i marken • Okänslighet för tillfälliga störningar • Temperaturutjämning • Smakförbättring (Karlstad kommun 2005) • Mikrobiologisk barriär Nackdelarna är att det är svårt att kontrollera grund­ vattnets väg från infiltrationen till vattenpumparna. Det finns risk för att tillfälliga vattenkvalitetsförsämringar påverkar det naturliga grundvattnet för lång tid om det infiltrerade vattnet inte kontrolleras. Det är svårt att kontrollera de faktorer som påverkar reduktionen av mikroorganismer. 2.4 Igensättningar Reduktionen av organiskt material och mikroorgan­ ismer är till stor del beroende på hur utvecklad en biohud är i infiltrationsbassängerna. Detta diskuteras i avsnitt 6.4.5. Men ett problem med vattenförsörjningsanläggningar med konstgjord grundvattenbildning är igensättningar av infiltrationsytan, d.v.s. bassängerna, brunnarna eller marken, med resulter­ ande minskning av infiltrationshastigheten. Igen­ sättningar beror på fysiska, biologiska och kemiska processer. Fysiska processer är ackumulering av oorganiskt och organiskt material i infiltrationsvattnet, lera och siltpartiklar, alger och andra mikro­ organismer. Det fina materialet i marken kan genom infiltrationen röra sig nedåt och fastna där marken har en lägre permeabilitet och skapa ett ogenomträngligt lager. Biologisk igensättning omfattar ackumuler­ ingen av mikroorganismer i vattnet samt tillväxt av mikroorganismer på och i sanden och bildar en bio­ hud av biomassa som sätter igen porerna. Kemiska processer är flockning och sedimentering av kalcium­ karbonat, fosfat och andra ämnen (Bouwer 2002). 3 Patogener och indikatororganismer 3.1 Patogena organismer Parasitism är ett symbiotiskt förhållande där en av parterna lever av eller skadar den andra parten. Den organism som drar fördel av den andra kallas en para­ sit. Alla parasitära organismer och virus som orsakar en sjukdom hos värden är patogena organismer (Grek­ iska patho, sjukdom och gennan, framkalla). Av trad­ ition brukar ”parasiter” avse protozoer och helminter. Patogener som sprids med vatten består huvudsakligen av virus, bakterier och protozoer (Prescott 1999). De flesta av de vattenburna virala, bakteriella och protozoiska patogenerna är av fekalt ursprung och sprids genom fekal-oral väg. De flesta orsakar magsjukdomar och påverkan på hälsan kan sträcka sig från infektioner utan symtom, till lindrig diarré, eller till allvarlig sjukdom som kräver sjukvård. 13 går sönder och de inre delarna, DNAt eller RNAt, bryts ner. Flera hundra typer av virus har identifierats i avföring. När en levande cell har angripits av ett virus, förvandlas cellen snabbt till en grynig massa av virus som är redo att infektera nya celler (HVR 2005). Bakteriofager är virus som angriper bakterier, men har för övrigt samma egenskaper som virus som angriper eukaryoter (se i Figur 3‑1). De indelas efter vilken bakterie de angriper och kan användas för identifiering av bakterier eftersom de är mycket specifika (se avsnitt 3.3.1) (Thougaard, Varlund & Madsen 2001). 3.1.1 Virus Virus är enkla, ocellulära entiteter (s.k. virioner) som består av en eller flera molekyler av DNA eller RNA. Dessa är inkapslade i ett proteinhölje, en kapsid, och ibland även flera lager med kolhydrater eller lipider, ett yttermembran. Proteinkapsiden består av joniser­ bara aminosyror (glutamat, aspartam, histidin, tyrosin). Liksom många organiska föreningar kan dessa, beroende på pH omkring föreningen, avge eller ta upp en elektron och därigenom få en laddning. Virus har därför varierande laddning och varje virus har en specifik isoelektrisk punkt där dess nettoladdning är noll (Zhuang & Jin 2003) (se även avsnitt 6.1.4). Virus är i storleksordningen 0,02–0,09 µm (20– 90 nm) och därmed mycket mindre än patogena bakterier och protozoer. De reproducerar sig endast inom andra levande celler och är uteslutande intracellulära parasiter. Virus skiljer sig från levande celler på tre sätt; deras enkla ocellulära uppbyggnad, frånvaro av både RNA och DNA i samma virion, samt förmågan att reproducera sig utan en värdcell. De markerar gränsen mellan levande organism och död materia och är inaktiva när de är utanför celler. Virus klassificeras genom deras morfologi; nukleinsyratyp (DNA eller RNA, enkelsträngad eller dubbelsträngad), kapsidens uppbyggnad och storlek, om viruset har ett yttermembran, samt vilka värdar de använder sig av och vilka sjukdomar de orsakar (Prescott 1999). Eftersom virus inte anses vara levande kallas avdödningen istället för inaktivering. Inaktivering av virus är en process som innebär att viruset förlorar förmågan att infektera dess värdorganismer, kapsiden 3.1.2 Bakterier Bakterier är encelliga organismer utan cellkärna och är den mest spridda av alla livsformer. Antalet arter uppskattas till 20–30 miljoner, men av dessa finns endast ca 5 000 fullständigt beskrivna. De flesta är helt ofarliga och hjälper oss t.ex. ta tillvara på vår föda på rätt sätt. Bakterier förekommer rikligt i de flesta miljöer och förökar sig genom delning. De spelar en mycket viktig roll i den kemiska omsättningen i naturen men kan också framkalla sjukdomar, dels genom bildning av gifter, toxiner, dels genom att reta den insjuknandes immunförsvar. (Wikipedia 2005). De patogena bakterierna varierar i storlek mellan ca 0,4–14 µm (400–1400 nm) i längd till 0,2–12 µm (200–1200 nm) i bredd (EPA 1993). Figur 3‑1. Virioners form och uppbyggnad. a och c är ikosaedriska virioner med resp. utan ett yttermembran, b och d är en helixformad virion och e är en bakteriofag med den karakteristiska svansen (Thougaard, Varlund & Madsen 2001, s. 149). 14 parvum. Salmonella och Shigella har förekommit i några enstaka fall vid smitta från enskilda vattentäkter/brunnar. Andra smittämnen som kan vara aktuella vid vattenburna utbrott i Sverige är: Yersinia enterocolitica, Aeromonas hydrophila, patogena E. coli (t.ex. enterohemorragisk E. coli, enteroaggreg­ ativa E. coli), hepatit A, rota-, coxsackie- och echo­ virus (tillhör familjen enterovirus) (Smittskydds­ institutet 2005). Helminter är idag inte intressanta ur hälsosynpunkt i svensk miljö (Rydberg 2005). Förekomst av patogener och indikatororganismer i yt- och grundvatten är beroende av en mängd fakt­ orer, bl.a. naturliga egenskaper hos avrinningsområdet och mänsklig påverkan som bidrar till utsläpp av patogener. Till ytvatten kan utsläppen vara punkt­ källor; avloppsreningsverk, eller diffusa; förorenad avrinning från jordbruksland eller djurhållning. Många patogener reduceras i ytvattnet genom utspädning, sedimentering och avdödning pga. att den naturliga miljöns förhållanden (temperatur, solljus och predation) skiljer sig från optimala förhållanden för överlevnad. Grundvatten är i normala fall mer skyddat från förorening än ytvatten genom barriäreffekterna som marken och dess omättade zon utgör. Förorenat grundvatten återfinns därför oftast där dessa barriärer är svagare och direkt kontakt mellan förorening och grundvatten kan ske (WHO 2004). Tabell 3-2 redovisar uppskattningar av höga kon­ centrationer av enteriska patogener och indikatororganismer i olika typer av ytvatten och grundvatten (taget från litteratur uppgifter) (WHO 2004). Den ger en indikation om vilka halter som kan tänkas uppkomma och ger en fingervisning av halter som kan jämföras med reduktionen som uppmätts vid infiltration av ytvatten i syfte att producera dricksvatten genom konstgjord grundvattenbildning som kommer redovisas i följande kapitel i litteraturstudien. Det finns dock ett antal begränsningar och osäkerhet i datan i tabellen, var halterna är uppmätta i förhåll­ ande till källan och känsligheten i analyserna. Det är stor skillnad mellan medelvärden som WHO redo­ visar och medelvärden för svenska sjöar och vattendrag, vilka är mycket lägre. I Sverige inkluderas inte virus av dricksvattenföre­ skrifterna. Ett allmänt angreppssätt som används för att beräkna och hantera risker med vattenburna pato­ gener är att utgå ifrån en maximal acceptabel risk för infektion av en på 10000 personer per år, vatten­ konsumtion och dos-respons relationer för olika 3.1.3 Protozoer Protozoer är encelliga organismer med en cellkärna (eukaryoter) och har vissa egenskaper som vanligtvis förknippas med djur, t.ex. förmågan att röra sig och heterotroft näringsupptag. Protozoer är större än bakterier och virus; 10–50 µm men det finns arter upp till 0,5 mm. De finns i stora mängder i vattenmiljöer och i marken och är viktiga för ekosystemet, eftersom de i huvudsak lever på bakterier. De delas upp i huvudgrupperna Flagellater, Amöbor, Ciliater och Spordjur, utifrån hur de förflyttar sig och skillnader i form och reproduktion (Wikipedia 2005; Prescott 1999; Thougaard, Varlund & Madsen 2001). Protozoer kan vara svåra att oskadliggöra med vanlig desinfektion. För att överleva i svåra miljöer kan visa arter utsöndra ett sekret som bildar ett skydd runt cellen och övergår i ett vilostadium, en s.k. ”cysta”. Amöbor kan i vattenledningar dessutom fungera som bärare av bakterier som normalt skulle ha dödats av klorering. Noggrann mekanisk filtrering är den säkraste barriären (HRV 2005). 3.2 Vattenburna patogener och lagstiftning En förteckning av vattenburna patogener är svår att få komplett. Det dyker ständigt upp nya typer och hur de infekterar är inte alltid känt. De potentiella vattenburna patogenerna inklud­ erar: • Virus, bakterier, protozoer och helminter/maskar som finns i Tabell 3-1 (förutom Schistosoma som främst sprids med infekterat ytvatten vid badning). • Helicobacter pylori, Tsukamurella som har fått ökad betydelse de senaste åren. • Isospora belli och microsporidia, för vilka det är troligt att de sprids med vatten, men inte säkerställt. • Bacillus, sprids med livsmedel, men det finns inte bevis för att de sprids med vatten. • Giftiga cyanobakterier (WHO 2004). De vanligaste anledning till utbrott i Sverige under senare år har varit Campylobacter, Giardia lamblia och norovirus, men även utbrott med toxinbildande E. coli, Entamoeba histolytica och Cryptosporidium 15 Tabell 3-1. Viktiga vattenburna patogener (WHO 2004). Patogen Betydelse för hälsan Överlevnad i vattenförsörjning Motståndskraft till klor Relativ infektivitet Burkholderia pseudomallei Liten Kan föröka sig Låg Låg Campylobacter jejuni C.coli Stor Medel Låg Medel Escherichia coli - Patogena stammar Stor Medel Låg Låg E. Coli - Enterohemorragiska Stor Medel Låg Hög Legionella spp. Stor Förökar sig Låg Medel Icke-tuberkulos mycobacteria Liten Förökar sig Hög Låg Pseudomonas aeruginosae Medel Kan föröka sig Medel Låg Salmonella typhi Stor Medel Låg Låg Andra salmonella Stor Kan föröka sig Låg Låg Shigella spp. Stor Kort Låg Medel Vibrio cholerae Stor Kort Låg Låg Yersinia enterocolitica Stor Lång Låg Låg Adenovirus Stor Lång Medel Hög Enterovirus Stor Lång Medel Hög Hepatit A Stor Lång Medel Hög Hepatit E Stor Lång Medel Hög Norovirus and Sapovirus Stor Lång Medel Hög Rotavirus Stor Lång Medel Hög Acanthamöba spp. Stor Lång Hög Hög Cryptosporidium parvum Stor Lång Hög Hög Cyclospora cayetanensis Stor Lång Hög Hög Entamöba histolytica Stor Medel Hög Hög Giardia intetinalis Stor Medel Hög Hög Naegleria fowleri Stor Kan föröka sig Hög Hög Toxoplasma gondii Stor Lång Hög Hög Dracunculus medinensis Stor Medel Medel Hög Schistosoma spp. Stor Kort Medel Hög Bakterier Virus Protozoer Helminter 16 Tabell 3-2. Exempel på höga koncentrationer (per liter) av enteriska patogener och fekala indikatorer i olika typer av vattentäkter (WHO 2004; Göteborgs VA-verk 2005; Strand 2005; Stockholm Vatten 2005). Patogen eller indikator Sjöar Campylobacter Salmonella Påverkade vattendrag Naturliga vattendrag Göta älv Lyckebyån, Mälaren Grund­ vatten (median 2004) Karlskrona 20–500 90–2500 0–1100 0–10 – 3–58000 1–4 – Neg E.coli 104 –106 3*104 –106 6000–3*104 0–103 980 580 30 Virus 1–10 30–60 0–3 0–2 4–290 2–480 2–240 0–1 2–30 1–470 1–2 0–1 Cl. perfringens 170 90 <10 Enterokocker 390 Cryptosporidium Giardia patogener. För virus fås en maximal tillåten koncentration på 1,8·10 –7 pfp/l baserat på beräkningar för rotavirus och poliovirus 3 (Schjiven et al. 2003). Det här tillvägagångssättet används bl.a. i USA och Nederländerna (Zuang & Jin 2003; Hijnen et al. 2004). Utgående från de uppmätta koncentration­ erna av virus i ytvatten (0,02–10 pfp/l) i Neder­ länderna krävs en reduktion av virus på 5–8 log10 för att producera ett säkert dricksvatten (Schjiven et al. 2003). Tyskland har dricksvattenföreskrifter ”Surface Water Treatment Rule” som säger att en 4 log avskiljning av virus krävs vilken typ av process det än är (Dizer et al. 2005). Nya Zeelands kvalitetskrav för dricksvatten för enteriska virus är <1 per 100 l och <1 E.coli per 100 ml (Pang et al. 2003). Dessa kvalitetskrav är inte lika restriktiva som i USA och Nederländerna vilket även avspeglar sig på erfordrat avstånd och uppehållstid för anläggningar med konstgjord infiltrat­ ion, se vidare kapitel 7. Fekal förorening kan påvisas genom att använda de flesta tarmorganismer som utsöndras av befolkningen. Kraven på en indikatororganism är att: • Den ska förekomma naturligt och hög halt i avföring från människor och djur. • Den ska inte föröka sig i naturliga miljöer, utan ha en avdödning i mark- och vattenmiljö som liknar de sjukdomsalstrare man önskar indik­ era. • Känsligheten för vattenreningsmetoder ska inte vara större eller avsevärt mindre än de sjukdomsalstrare man önskar indikera. • Kvantifiering i labb ska kunna vara enkel och säker, med bra analysmetoder (Stenström 1985). De organismer som används som indikatororganismer förekommer i olika utsträckning i fekalier från människa och djur, reagerar olika i miljön och skiljer sig i hur resistenta de är mot olika behandlingar. En uppdelning kan därför göras i en grupp som indikerar för fekal påverkan och en grupp av organismer som kan användas för att mäta en behandlings effektivitet. De flesta organismerna kan användas till båda uppgifterna men passar bättre för den ena eller den andra (WHO 2004). Tabell 3-3 ger exempel på använda indikatororganismer. Även fysikalisk – kemiska indikatorer (turbiditet, konduktivitet, färgtal, absorbans, pH, BOD/COD/ eller TOC) används för att påvisa möjlig fekal påverkan. Dessa är grova och okänsliga och kan inte med säkerhet visa på om vattnet är förorenat med patogener eller ej. Men det finns även fördelar med dem; de är enkla, billiga, snabba och vissa kan mätas on-line. De är ofta analyserade och har på det sättet blivit allmänt vedertagna. En kombination av flera indikatorer är oftast det bästa. 3.3 Indikatororganismer Eftersom flera olika faktorer påverkar förekomsten av patogener i mark och vatten, och det är svårt att ut­ föra kvantitativa rättvisande analyser av sjukdoms­alstr­ ande bakterier, virus och parasiter eftersom de före­ kommer i så låga koncentrationer, indikeras därför en risk indirekt med hjälp av förekomsten av normala tarmbakterier och andra indikatororgan­ismer. Dessa är inte sjukdomsframkallande men visar med sin närvaro att det finns fekal förorening och därmed risk för sjukdomsalstrande bakterier (Stenström 1985). 17 Tabell 3-3. Indikatororganismer (WHO 2004; Göteborgs va-verk 2004). Totala koliforma bakterier Finns i både naturliga vatten och avlopp. De kan även indikera fekal påverkan och förorening, men är främst indikatororganism för behandlingseffektivitet, de ska inte förekomma direkt efter en behandling. E.coli och termotoleranta koliforma bakterier Typ av koliform bakterie som finns i djur- och människoavföring, Indikator på fekal förorening. E.coli är känsligare för desinfektion än virus och protozoa, och har en relativ hög avdödning. Ej lämplig indikator för behandlingseffektivitet. Heterotrofa bakterier Indikator för behandlingseffektivitet och desinfektion, tillväxt i vattenverk eller rörnät. Enterokocker Indikator på fekal förorening, överlever längre i vatten än E.coli och är mer resistenta mot klorinering och uttorkning, ej lämplig indikator för behandlingseffektivitet. Clostridium perfringens Sporbildande bakterie som har hög överlevnad och motståndskraft, är därför lämplig som indikator på enteriska virus och protozoa, samt allmän fekal påverkan Kolifager Virus som använder E.colibakterier som värdar för sin reproduktion. (Se diskussion i avsnitt 6.3.1) Bacteroides fragilis fager Som ovan, men har B.fragilis som värd. Är mycket resistent mot ogynnsamma förhållanden. Indikator på fekal förorening, finns endast i människors avföring. (Se avsnitt 6.3.1) I tarmen utsöndras bl.a kolesterol som omvandlas av tarmmikroorganismer till ett flertal andra sterolföreningar (t.ex. coprostanol). Sterolföreningar kan detekteras kemiskt och är en indikator för fekal påverkan, som ger möjlighet att skilja på påverkan från människor eller betesdjur. Detta är en metod som är relativt ny och används inte som standardanalys i Sverige (Ottoson et al. 2003). inte föröka sig i naturlig miljö, uppträda i samma mängd eller mer än enteriska virus samt ha samma överlevnadsförmåga (Leclerc et al. 2000). Ett visst virus lämplighet som indikatororganism bestäms med andra ord av dess relativa okänslighet för in­ aktivering (Yates et al. 1985, citerad i Jin, Chu & Li 2000) och dess ekologiska och morfologiska likheter med virus som infekterar människor (Havelaar et al. 1993, citerad i Jin, Chu & Li 2000). Tre grupper av bakteriofager används huvudsakligen som indikator- och modellorganismer; Som­ atiska kolifager (fager som angriper bakteriecellens huvudkropp), FRNA fager (angriper cellen på F pili) och fager som infekterar bakterien Bacteroides fragilis. Problemet med att använda somatiska kolifager som fekala indikatororganismer är att de är för generella och använder flera bakterier som värdar för sin reproduktion, inte bara fekala koliformer. Leclerc m.fl. (2000) hävdar att vissa typer av fager kan infektera den normala bakteriefloran i mag-tarmkanalen och kan detekteras i avloppsvatten, men de finns sällan i mänskliga fekalier. Kolifager har både upptäckts i vatten utan fekal påverkan och har inte kunnat detekteras i vatten med kraftig fekal förorening enligt Leclerc m.fl. (2000). Fagerna är alltså indikatorer av indikatorer (E.coli och koliformer) som i sin tur måste finnas i en koncentration av minst 10 4/ml. Fagen är således inte lämplig som indikator för fekal förorening eller enteriska virus. 3.3.1 Bakteriofager som indikatororganismer och modellorganismer Liksom för bakterier finns det för många kända enteriska virus för att övervakning ska kunna ske utifrån dessa. Dessutom finns de endast i låga koncentrationer vilket gör analysen av virus svår. Efter­ som de ordinära indikatororganismerna skiljer sig från enteriska virus till storlek, resistens för vattenbehandling och överlevnadstid i naturen, kan dessa inte ge en fullgod bild av virus utbredning. Istället kan bakteriofager (el. fager); virus som angriper bakt­ erier, användas som indikatororganismer och modell­ organismer (Leclerc et al. 2000) Som för de redovisade indikatororganismerna i avsnitt 3.3 gäller samma villkor för fagerna om de ska vara tillfredställande indikatororganismer. De ska uppträda explicit och genomgående i avloppsvatten, 18 Den andra gruppen, FRNA bakteriofager, utsöndras i stor mängd i fekalier och finns i koncentrationer upp till 106 pfu/100 ml i obehandlat avloppsvatten (Pang et al. 2003; Leclerc et al. 2000). FRNA fager är enligt Nasser m.fl. (1993, citerad i Pang et al. 2003) mycket tåliga och mycket resistenta till in­ aktivering. Överlevnadsförsök i laboratoriemiljö med FRNA fager visade ingen inaktivering efter 8 dagar i 10 °C grundvatten spikat med avlopp (Pang et al. 1997, citerad i Pang et al. 2005) och inaktiveringshastigheten för MS2 var lägre än för poliovirus och echovirus vid samma temperatur. Den sista gruppen fager som använder Bacteroides fragilis som värd, är mycket mer resistent mot vatten­ behandling än de andra två grupperna och är lika persistent i naturlig miljö som FRNA fager och finns dessutom i stor mängd i mänskliga fekalier (Leclerc et al. 2000). Det finns alltså tveksamheter och problem med att använda bakteriofager för indikatorsyfte. Låg koncentration av alla typer av bakteriofager i grund­ vatten begränsar deras förmåga att agera som indik­ atororganismer för enteriska virus (Leclerc et al. 2000). Däremot är de mycket användbara som modellorganismer av virus i undersökningar av virus trans­ port och reduktion i mark och andra beredningsprocesser inom dricksvattenförsörjning. Bakterio­ fagerna går att tillsätta i höga koncentrationer vid försök, vilket gör det möjligt att undersöka redukt­ ionen. Det skulle inte vara praktiskt möjligt eller tillåtet att tillsätta virus till ett infiltrationssystem som är i drift. Vidare är analysmetoderna för fager är enklare och billigare än metoderna för enteriska virus (Peters et al. 1998) Eftersom virus egenskaper är varierande, och därmed dess transport och fastläggning i marken, är det inte troligt att endast en bakteriofag kan representera den stora mängden av enteriska virus som finns (Jin, Chu & Li 2000). Därför borde ett konservativt angreppssätt väljas med en eller flera bakt­ eriofager med olika egenskaper och helst med sämre avskiljning än de virus som önskas renas. De bakteriofager som är mest använda som simul­ erings/modell/surrogatorganismer i försök är MS2, PRDI, φX-174 och H40/1. MS2 är en enkelsträngad RNA bakteriofag utan yttermembran med 31 % innehåll av nukleinsyra. Den har en diameter på mellan 24–26 nm, vilket ligger i samma storleksordning som viktiga enteriska virus. φX-174 är en somatisk enkelsträngad DNA bakteriofag med ett innehåll av nukleinsyra på 26 % och med en diameter på mellan 25–27 nm. Deras isoelektriska punkt är 3,9 resp. 6,6 (Chattopadhyay & Puls 1999; Zhuang & Jin 2003). PRD1 är en ikosaedrisk fag med en diameter på 62 nm och med ett inre lipodmembran som gör den hydrofob. Dess pHiep ligger på mellan 3–4. H40/1 är en marin bakteriofag som infekterar en icke-patogen bakterie, Pseudoalteromonas gracilis, som normalt inte existerar i grundvatten. Fagen är 39 nm i diameter med en 46 nm lång svans och har en pHiep på 5,6 och är hydrofil (Flynn, Rossi & Hunkeler 2004). I allmänhet ger MS2 och FRNA fager en konservativ bild av transporten och rörelsen hos virus pga. den relativt dåliga fastläggningen vid negativt laddade jordpartiklar och pH 6–8, jämfört med andra enteriska virus (Jörgensen, Peters & Eschweiler 2001). φX-174 används även den som simuleringsorganism, men oftast som en jämförelse med MS2 bl.a. för att kunna studera skillnaden i adsorption pga. de olika pHiep. φX-174 har vid naturliga pH bättre fastläggning än de andra beskrivna fagerna (Schjiven, Hassanizadeh & de Bruin 2002). An­ vändning av Bacteroides fragilis fager som modellorganismer är begränsad pga. att värdbakterien är svår att odla då den är anaerob (Leclerc et al. 2000). 4 Studier om mikrobio­ logisk barriärverkan vid konstgjord grundvatten­ bildning Mikrobiologisk barriärverkan vid konstgjord grund­ vattenbildning, inaktivering, transport och fastläggning kan studeras på flera sätt och syftet med studien bestämmer metoden. De metoder som används är i olika skala; satsvisa försök, kolonn eller fullskaleförsök i fält. I satsvisa försök är det i huvudsak jämvikts adsorption och inaktivering/avdödning som studeras, och dispersion kan inte undersökas. Kolonnstudier blir mer lika verkliga förhållanden, men har fortfarande flera svagheter jämfört med fältstudier, bl.a. korta avstånd och uppehållstider. 19 Kolonnstudier kan innefatta dispersion om materialet inte är homogent packat. I kolonner som är packade uniformt kan dispersionen försummas. I fältstudier undersöks de riktiga förhållandena ofta genom tillsats av spårämnen och simuleringsorganismer (bakt­ eriofager), s.k. spikning. Beroende på den hydrolog­ iska situationen kan transporten av mikroorganismer modelleras som 1-, 2-, eller 3-dimensionell. Dispers­ ion kan vara en betydande effekt vid verkliga förhåll­ anden (Schijven 2001). Satsvisa- och kolonnförsök är ofta mycket specialiserade för vissa förhållanden, t.ex. en viss sand och indikatororganism, vid olika pH. För fältstudierna är det fler faktorer som påverkar och det är mycket sällan som patogena organismer används i studierna, utan nästan uteslutande modellorganismer såsom bakteriofager. Mycket varierande resultat har erhållits från dessa undersökningar beroende på förutsättningarna, vilket beskrivs i kapitel 7. För denna litteraturstudie har i huvudsak artiklar om satsvisa försök och kolonnförsök analyserats för att bedöma de olika faktorernas påverkan (även andra litteraturstudier och sammanfattningar har använts). Beskrivningar av fältstudier har använts för kvantifieringsavsnittet, eftersom de utfördes under naturliga förhållanden och är någorlunda jämförbara. 5 Påverkande faktorer 5.1 Översikt Två övergripande principer påverkar den mikrobiologiska barriärverkan; inaktivering och avskiljning. Helst ska bägge principerna inkluderas vid beredning av dricksvatten, vilket de gör vid konstgjord grundvattenbildning. Olika faktorer påverkar mekanism­ ernas effektivitet och detta ska granskas i följande kapitel. Det är skillnad på mekanismerna mellan bakt­ eriers, parasiters och virus reduktion, och därför kan olika parametrar som påverkar inaktivering och avskiljning påverka mikroorganismerna på olika sätt (Stenström 1980). Begreppen i Figur 5-1 kan definieras och relateras till varandra på lite olika sätt. Reduktion mäts oftast som andel mikroorganismer som passerar en process och redovisas i logaritmisk skala (–log10 (C/C 0)). Reduktionen kan även behandlas och modelleras som t.ex. kollisionseffektiv­ itet, relationen mellan fastläggning och lossning m.m. beroende på vilken modell som används. Avskiljningen kan ske genom filtrering och silning. Enligt Foppen, Mporokoso och Schijven (2005) och Stevik m.fl. (2004) är filtrering borttagning av part­ ikelmassa från en lösning genom kollisioner med och deposition på ytan på materialet i det porösa mediet. Termen inkluderar både sedimentation, kinetisk- och jämviktsadsorption, samt hydrofoba interaktioner. Mikrobiologisk barriärverkan - effektiviteten mäts i reduktionen av mikroorganismer Inaktivering (Virus) Avdödning (Bakterier och protozoer) Avskiljning/Fastläggning/Retention kan ske genom: Silning (mekanisk filtrering), grövre partiklar Filtrering, finare partiklar - Adsorption - Hydrofob interaktion Figur 5-1. Begrepp inom mikrobiologisk barriärverkan. 20 En jämvikt ställer in sig mellan koncentrationen av mikroorganismer i lösningen och på materialet under satsvisa- och kolonnförsök. Detta betecknas som jämviktsadsorption, vilket egentligen inte bidrar till reduktionen av mikroorganismer. Den kinetiska ad­ sorptionen är det som till stor del kontrollerar reduktionen och verkar innan ett jämviktstillstånd uppstår. Även den består av fastläggning och lossnande från materialet, men fastläggningen är så mycket snabbare och mer betydande än frisättningen av de adsorberade mikroorganismerna. Hydrofob interaktion resulterar i en ökning av fastläggningen genom att hydrofoba grupper på ytan på mikroorgan­ ismen och på det fasta materialet interagerar (Schijven 2001, s.28). Silning (eng. straining) är infångandet av mikroorganismer i små porer som de inte kan passera, även kallad mekanisk filtrering. Om silning och/eller filtr­ ering sker beror av storleken på materialet och mikro­ organismen. I vissa äldre studier (bl.a. Stenström 1980, 1985) används endast filtrering som namn på det som kallas silning eller mekanisk filtrering i denna studie. Mikroorganismers beteende i grundvatten och vid infiltration kontrolleras av egenskaperna hos mikroorganismen själv och egenskaperna hos vattnet och marken (Guan et al. 2003) se figur 5-2. 5.2 Avskiljning Den mikrobiologiska barriären vid infiltration av ytvatten styrs förutom av mikroorganismernas inaktivering och avdödning även av avskiljning genom filtrering och/eller silning. Filtrering, som inkluderar adsorption, är en process som gör att patogener stannar på ytor i akvifären och kan vara reversibel eller irreversibel. Omständig­ heter som påverkar adsorptionen av virus till jordpartiklar inkluderar elektrostatisk attraktion och repulsion, van der Waals krafter, kovalenta- och jon­ bindningar och hydrofobiska interaktioner. Dessa påverkas i sin tur av pH, innehåll av organiskt materi­ al, järn och andra joner, vilka har visats sig mycket betydelsefulla vid adsorptionen (Jin et al. 1997). Virus påverkas i högre grad av fysikalisk-kemiska adsorptionsprocesser än bakterier i marken (Stenström 1980). Egenskaper hos mikroorganismen Koncentration Organismtyp Storlek och form Laddning Hydrofobicitet Egenskaper hos vattnet pH Temperatur Innehåll av organiskt material Jonkoncentration och jonsammansättning Mikrobiologisk barriärverkan Egenskaper hos marken och infiltrationsmaterialet Partikel- och porstorlek Mineral- och jordsammansättning Laddning på markpartiklarna Hydrofobicitet Koncentration i grundvatten Anläggningens utformning och drift Uppehållstid Transportsträcka mellan infiltration och uttag Grundvattenhastighet Vattenmättnadsgrad Biohud och naturliga mikroorganismer Figur 5-2. Faktorer som påverkar mikroorganismers överlevnad och avskiljning i marken (fritt efter Stenström 1985). 21 För bakterier och protozoer är adsorptionen en viktig faktor, men här inverkar även silningseffekter. Den passiva transporten av bakterier med infiltrerande vatten är mer betydande än diffusion och aktiv rör­ else av bakterien (Jörgensen, Peters & Eschweiler 2001). Storleken på partiklarna i infiltrationsmateri­ alet och bakteriernas storlek, samt den hydrauliska belastningen och igensättningar, påverkar silningen. Även adsorption av bakterier påverkas av innehållet av organiskt material, till vilken utsträckning en bio­ film utvecklas och elektrostatisk attraktion mellan laddningar på cellens eller partiklarnas yta (Stevik et al. 2004). Adsorption är det begrepp som används mest frekvent i studier om reduktion av virus. Ab­ sorption kan även ske till materialet om det förekommer ett lager av organiskt material/biofilm runt partiklarna, och därför kan sorption vara det begrepp som är riktigt att använda om osäkerhet föreligger om vilken process som styr reduktionen, se Figur 5-3. och joninnehåll samt en konstant låg temperatur. Den biologiska aktiviteten och predationen är låg och detta gör att organismer som når grundvattnet kommer att överleva under en längre tid. Över­ levnadsgraden skiljer sig mellan olika organismer, men även mellan stammar inom en bakterieart kan det vara skillnad (Stenström 1985). Inaktivering av virus sker om temperaturen höjs till 50–60 °C i ca 30 minuter, att pH sänks till <5 eller genom bestrålning, vilket får betraktas som ganska extrema betingelser. De är dessutom mycket resistenta mot desinfektion med klor och klarar 3 ggr högre klorkoncentration än bakterier (Thougaard, Varlund & Madsen 2001). Eftersom inga av dessa förutsättningar existerar ute i naturliga miljöer påverkas virus inte avsevärt av inaktivering vid konstgjord grundvattenbildning. 5.4 Relationen mellan inaktivering och avskiljning Virusadsorption till markpartiklar tycks vara den mest betydelsefulla processen för reduktion av virus. Men för att åstadkomma en fullständig borttagning av virus från en lösning måste det ske genom avdödning och irreversibel adsorption. Under de försök som studerar avskiljning av virus antas avdödningen vara försumbar under de förhållanden som råder och den korta tid som experimenten pågår (Schjiven et al. 2001). En jämförelse i en modellering av Schjiven m.fl. (2001) mellan inaktivering och adsorption av data från flera fältstudier, visar att adsorption är av större betydelse än inaktivering vid infiltration. Pang m.fl. (2003) utförde kolonn- och satsvisa experiment för att bestämma det relativa bidraget från avdödning och överlevnad till reduktionen. Av­ skiljningen av bakterier skedde främst genom filtrer­ ing (87–88 %) och delvis genom avdödning (12– 13 %), medan virusavskiljning var ungefär till lika delar filtrering (55 %), och inaktivering (45 %). Att studien fick ett resultat som skiljer sig till viss del från teorin ovan kan bero på att materialet var mycket fint och en stor andel bakterier filtrerades bort, vilket inte skedde till lika stor utsträckning för fagerna. En viss tillväxt av bakterier skedde och avdödningen för bakterier blev därmed långsammare än inaktiveringen för fagerna. Försöken utfördes i rumstemperatur och Figur 5-3. Adsorption – olika begrepp (Ederth 2005). 5.3 Inaktivering och avdödning Miljön i marken skiljer sig signifikant från den miljö i kroppen som patogener normalt existerar i. Det be­ tyder att de så småningom dör av, men överlevnadstiden kan vara lång. Hastigheten varmed de patogena mikroorganismerna dör påverkas av både abiotiska (icke levande) faktorer, t.ex. vatteninnehåll i marken, pH, temperatur, mängden organiskt material och bio­ tiska (levande) faktorer; organismart och predation och antagonistisk symbios mellan mikroorgan­ism­ erna i systemet (Stevik et al. 2004). Men överlag re­ presenterar grundvattnet en miljö med utmärkta för­ hållanden för mikroorganismers överlevnad. Miljön är stabil med små fluktuationer av vattenhalt, pH fientlig samlevnad, parasitism. 22 avdödningen kan då vara relativt hög. Resultaten är därmed inte allmängiltiga utan det relativa bidraget från avskiljning och inaktivering tycks enligt studien variera beroende på rådande förhållanden. 6.1.2 Organismtyp Olika mikroorganismers beteende i en miljö med naturligt material styrs till stor del av laddningen på virusets yta och dess hydrofobicitet som kan skilja sig mycket mellan olika typer och även inom samma art, se följande avsnitt. I försök som varit väldefini­ erade och under exakt samma förhållanden, har olika beteenden observerats även inom samma stam av virus (Schjiven et al. 2001). Reduktionen blir därmed beroende av vilken typ av mikroorganism det handlar om. I studier väljs därför överlag olika mikroorgan­ ismer och helst s.k. konservativa indikatorer, för att få en så bra bild som möjligt av reduktionen och inte överskatta den mikrobiologiska barriären. 6 De olika faktorernas inverkan på reduktionen av patogener Detta kapitel beskriver faktorerna i figur 5-2 mer ingående. 6.1.3 Storlek och form Storleken på patogenen påverkar silning, filtrering och adsorption. Filtreringseffekten är liten för enskilda bakterier och virus enligt Stenström (1985), som skriver att storleken är av underordnad betydelse eftersom filtrering inte är en betydande reduktionsmekanism. Ju större en organism är desto större inverkan får filtreringseffekten och för parasiter är den mer betydande. Risken för parasitspridning an­ ses mindre i grundvatten av denna orsak (Stenström 1980). Storleken har däremot visat sig ha betydelse i andra studier, eftersom större mikroorganismer har färre porer att ta sig igenom och därmed silas bort. Flera studier visar att bakterier som är långa och ovala avskiljs till större grad än mindre bakterier. Detta gäller speciellt vid omättade förhållanden, efter­ som de större porerna är de första att torka ut. Om cellstorleken på bakterierna minskar pga. näringsbrist resulterar det i att bakterierna kan penetrera djupare (Stevik et al. 2004). Virus anses kunna transporteras längre i marken och penetrera djupare, just av den anledningen att de är mindre än ca en tiondel av storleken mot bakterier och protozoer (Macler & Merkle 2000). För virus resonerar Chattopadhyay och Puls (1999) att ju större ytarea ett virus har, desto fler adsorptions­ platser på kapsiden eller ytterhöljet och fler chanser till interaktion med adsorbanten. Chansen för ett 6.1 Egenskaper hos mikroorganismen 6.1.1 Koncentration Foppen, Mporokoso och Schijven (2005) undersökte hur filtrering i små porer, som håller kvar mikroorgan­ ismer, påverkades av koncentrationen av mikroorgan­ ismer i det infiltrerade vattnet. Försöket genomfördes i kolonner och genombrottskurvorna visade att vol­ ymen av vatten som krävdes för att fylla de små porerna med mikroorganismer var proportionell mot koncentrationen av bakterier i vattnet. Efter att por­ erna fyllts med mikroorganismer blev deras bidrag till reduktionen mindre och genombrottet större, d.v.s. reduktionen minskade. Vid en hög utgångshalt försämras därför reduktionen snabbare än vid en låg halt. Denna teori styrker Stevik m.fl. (2004) som skriver att försök genomförts som visat att en ökning av bakteriekoncentrationen ökar kollisioner med materialet och därigenom möjligheter till adsorption. Adsorptionshastigheten antas öka linjärt med bakteriekoncentrationen tills ett mättat tillstånd uppstår i materialet. Stenström greppet. 23 (1985) inkluderar inte adsorption i filtreringsbe- virus att bli påverkad av elektrostatiska eller hydrofoba krafter är direkt proportionell mot ytarean, stor­ leken har därför betydelse även för virusreduktion. Ytkaraktären hos bakterier, t.ex. om de har ett flagellum eller utskott, kan påverka fastläggningen positiv (Stevik et al. 2004). Om bakterien har en mobil förmåga kan detta också öka adsorptionen. 6.1.5 Hydrofobicitet Hydrofobicitet är en partikels vattenavstötande förmåga och en hydrofob partikel är vattenfrånstötande. Hydrofoba mikroorganismer är ofta elektriskt neutr­ ala och icke-polära, och föredrar därför neutrala och icke-polära lösningar eller miljöer. Adsorptionen är därigenom bättre för en hydrofob mikroorganism till en hydrofob partikel än en hydrofil partikel. Hydro­ fobiciteten bestämmer den hydrofoba interaktionen med adsorbanten och verkar vara en starkare kraft än ytladdning och dominerar därför för hydrofoba organismer. (Stevik et al. 2004). Hydrofobiciteten hos en mikroorganism eller partikel är proportionell mot kontaktvinkeln som bildas mellan vattnet och mikroorganismen (Chattopadhyay & Puls 1999). Chattopadhyay & Puls (1999) hävdar att det är de hydrofobiska krafterna som styr adsorptionen av virus till jord. Studien genomfördes som satsvisa försök med bakteriofagerna MS2, φX-174 och T-2 till tre typer av mineral och en lera. En termodynamisk studie utfördes för de olika adsorptionsexperimenten för att bestämma processens förmåga att genomföras och den relativa styrkan hos olika krafter genom att mäta den totala fria energin (∆G) vid interaktionen mellan de olika partiklarna. ∆G antogs vara summan av ∆GH (från hydrofobiska interaktioner) och ∆GEL (från elektrostatiska interaktioner). Det visade sig att under de förhållanden som undersöktes, var hydrofobisk adsorption mer betydelsefull än elektrostatisk adsorption, då de elektrostatiska krafterna endast utgjorde en liten del av den totala förändringen i fri energi. För de olika fagerna var adsorptionen störst för T-2 (93–97 % beroende på lera), MS2 (92–94 %) och minst för φX-174 (88–93 %). Detta stämmer med fagernas olika kontaktvinklar med vatten som var i ordningen T-2>MS2>φX-174. 6.1.4 Laddning hos mikroorganismerna Laddningen på mikroorganismen och viruset bestämmer den elektrostatiska interaktionen med en adsorbent. Laddningen på virusets ytterhölje är beroende av pH-värdet i lösningen (se även 6.2.1 pH). Isoelektrisk punkt är det pH-värde för vilket ett proteins nettoladdning är 0 och benämns pHiep, pHisp, IEP eller pI. Detta påverkar virus, som tidigare nämnts i avsnitt 3.1.1, eftersom dess yttermembran, kapsiden, är uppbyggd av proteiner. Virus får olika isoelektrisk punkt beroende på hur de är uppbyggda. Proteinet är som minst lösligt i vatten vid sin isoelektriska punkt. Virus blir därmed mer eller mindre laddade och vattenlösliga vid olika pH, beroende på vilken typ av virus det gäller (Guan et al. 2003; Jin et al. 1997). En partikel är negativt laddad om pH är över dess pHiep och vice versa. I kolonnförsök som utfördes i korta mättade kolonner på 20 cm adsorberades inte bakteriofagen MS-2, men en betydande mängd φX-174, ca 80 %, fastlades i kolonnerna. Detta berodde förmodligen på fagernas skillnad i isoelektrisk punkt. Både MS2 och sanden hade låga pHiep (3,9 resp 2–3) och var följaktligen negativt laddade vid experimentets pH 7,5 och repellerade varandra. För φX-174 var retent­ ionen större än för MS2 pga. att φX-174 har en högre pHiep (6,6) och dess negativa laddning var svagare och repulsionen därmed inte lika kraftig mellan fagen och materialet (Jin et al. 1997). Elektrostatiska laddningar påverkar även bakterier och andra mikroorganismer på liknande sätt som för virus. Laddningarna på en mikrobiell yta uppkommer genom dissociation av olika oorganiska grupper, t.ex. karboxyl- och aminogrupper lokaliserade på ytan eller djupare in i cellväggen som förändras vid olika pH. För de flesta bakterier ligger isoelektriska punkten mellan pH 1,5 och 4 (enligt Jörgensen m.fl. (2001) mellan 2,5 och 3,5) och de är negativt laddade i den naturliga miljön (Stevik et al. 2004). 6.2 Egenskaper hos vattnet 6.2.1 pH En mängd motstridiga resultat finns redovisade vad gäller pHs påverkan på adsorptionen av mikroorgan­ ismer vid olika försök. Förmodligen beror detta på olika isoelektriska punkter och skillnader i laddningar runt den isoelektriska punkten för olika arter 24 (Stevik et al. 2004). Men allmänt vedertaget är att pH spelar en viktig roll för mikroorganismernas ad­ sorptionsprocesser i mark, eftersom pH påverkar ladd­ ningen på mikroorganismerna som ska adsorberas och på det permeabla materialet som är adsorbanten (se även avsnitt 6.1.4 Laddning hos mikroorganismen och avsnitt 6.3.3 Laddning hos mark- och infiltrat­ ionsmaterialet). Tecknet på laddningarna kan bli det omvända be­ roende på förhållandet mellan de isoelektriska punkt­ erna för partiklarna och vattnets pH. Om pH ligger så att laddningarna är motsatta, är fastläggningen bättre. En partikel är negativt laddad om pH är över dess pHiep och vice versa. Detta medför att attrakt­ ionen mellan partiklar med olika pHiep kan uppvisa en dramatisk förändring vid ett specifikt pH som orsakar motsatt laddning hos dem. Guan m.fl. (2003) visar ett sådant beteende för fagerna MS2 och φX-174 vars avskiljning studerades i kolonner vid olika pH. När pH i lösningen var under 5 adsorberades MS2 irreversibelt på mineralen fältspat som var positivt laddad vid detta pH och mycket sämre över pH 5. φX-174 adsorberades bäst vid ett pH under 6, fagen var positivt laddad men mineralen kvarts negativt laddad. Det pH vid vilken adsorptionen förändras dramatiskt, det ”kritiska pH” ligger enligt Guan m.fl. (2003) ungefär en halv pH enhet under den högsta av virusets eller adsorbentens pHiep. Teorin om ett kritiskt pH borde även gälla andra mikroorganismer, men bakterier har överlag lägre pHiep (1,5–4) än virus och pH blir därmed inte en lika viktig faktor eftersom bakterier inte påverkas på samma sätt av detta fenomen i naturliga miljöer (Jörgensen et al. 2001). Även om pH ligger på ett värde där adsorption inte är gynnsam p.g.a. repulsion, sker ändå en fastläggning av virus eftersom andra processer verkar; van der Waals krafter, hydrofobiska effekter och filtrering. Den elektrostatiska repulsionen kan hindra effekten av dessa processer, men inte utesluta dem (Guan et al. 2003). Enligt Leclerc m.fl. (2000) ökar generellt adsorpt­ ionen av virus när pH minskar, men det finns lite olika trender beroende på komplicerade förhållanden. Enligt Schjiven (2001) beror detta på den ökade elektrostatiska repulsionen vid högre pH (både viruset och materialets negativa laddning ökar överlag med ökat pH) och Figur 6-1 visar koefficienter för fastläggning/lossnande i en modell, för olika grupper av virus vid olika pH. För överlevnaden av mikroorganismer spelar pH en liten roll, men vid riktig låga och höga värden får det en inverkan. Överlevnaden är överlag bättre vid lite högre pH än i sura jordar (Stevik et al. 2004). Vid pH >10 denaturerar virusets ytterhölje eftersom det består av en proteinstruktur. 6.2.2 Temperatur För adsorption av bakterier gäller att vid en högre temperatur är fastläggningen bättre vilket kan bero på en ökning av viskositeten av vattnet, bakteriens yta samt förändringar i fysiologin hos organismerna (Stevik et al. 2004). Överlevnaden för mikroorganismer i mark och vatten är däremot i regel längre vid en lägre temp­eratur. Vid 4 °C är enligt Stenström (1985) överlevnadstiden Figur 6-1. Medelvärden på fastläggning/lossnande (katt/kdet) i relation till pH för två virusgrupper (Grupp I: MS2, φX-174, Eckovirus, CB4 m.fl. Grupp II: T-2, T-4, Poliovirus, CB3 m.fl.) (Goyle och Gerba 1979 refererade i Schjiven 2001). 25 för mikroorganismer upp till 6 månader om övriga betingelser är goda. Men en lägre temperatur bidrar även till trögare vatten och längre uppehållstider, vilket till viss del motverkar effekten av längre överlevnad. En högre temperatur bidrar även till högre aktivitet hos predatorer, vilket gör att överlevnaden minskar (Stenström 1980). Temperaturen anses vara den dominerande och mest betydande faktorn för virus inaktivering i vatten och mark (Jin et al. 1997). Temperaturen är den enda signifikant korrelerade parametern till inaktiver­ ing av virus i grundvatten och samma observation har gjorts för virus i marken (Yates et al. (1985) citerad i Jörgensen et al. 2001). Detta diskuterar i Rzezutka och Cook (2004) i sin genomgång av studier på inaktivering av virus i vatten, mark, mat och på andra fasta ytor. Temperaturen är av stor betydelse men den har även en stor inverkan på andra mikroorganismer som i sin tur påverkar inaktiveringen av virus. Schjiven (2001) har sammanställt avdödningskoefficienter för virus i grundvatten från olika studier och ett samband syns i Figur 6-2. Studierna tyder således på att hög temperatur ger bättre fastläggning och högre avdödning. organiskt material som har liknande pH iep som mikroorganismerna (Jörgensen et al. 2001; Leclerc et al. 2000). Efter regn finns tydliga indikationer på att virus desorberas, vilket kanske kan bero på en ökad mängd organiskt material som infiltreras och ett utbyte sker på sedimentens yta (Stenström 1985). Enligt Stevik m.fl. (2004) skiljer sig resultatet av hög halt av organiskt material beroende på om det sitter på materialet eller finns i vattnet. Om det sitter på det fasta materialet kan det öka katjonbyteskapaciteten, ytarean och antalet ”sites”/platser för adsorption. Om det är en hög halt av organiskt material i vattnet kan det minska adsorptionen för att det konkurrerar med mikroorganismerna om adsorptionsplatser. Pang m.fl. (2005) gjorde en modell av de infiltrat­ ionsavstånd som krävs för fullgod avskiljning enligt kvalitetskrav på dricksvatten i Nya Zeeland vid olika akvifärer. Det visade sig att ett kortare avstånd behövdes då akvifären var ren och inte hade blivit kon­ taminerad av organiskt material från tidigare utsläpp. Hade marken använts till avloppsinfiltration krävdes ett längre avstånd för att få fullgod rening, troligen pga. att det organiska materialet blockerade de bind­ ningsplatser som annars hade bundit patogener (Pang et al. 2005). Ju högre halt organiskt material desto längre överlevnad för bakterier och parasiter eftersom tillgången på näringsämnen är större (Stevik et al. 2004). Detta gäller även för virus; det organiska materialet skyddar från inaktivering. Men det organiska materi­ alet gynnar tillväxten av naturliga mikroorganismer som har en antagonistisk effekt på den patogena organismen (Stenström 1980). Om halten av suspenderat material är hög ökar filtreringseffekten, eftersom även biohuden påverkas och reducering av bakterier och protozoer blir effektivare. 6.2.3 Innehåll av organiskt material Det finns lite olika teorier om hur organiskt material i infiltrationsvattnet påverkar reduktionen av patogener. Det organiska materialet påverkar både adsorption och inaktivering av mikroorganismerna. En hög koncentration av organiskt material och ytaktiva ämnen minskar adsorptionen av mikroorgan­ ismer. Detta beror förmodligen på konkurrens om de hydrofoba adsorptionsplatser på sedimenten från Figur 6-2. Inaktiveringskoefficient för olika virus i lösning relaterad till temperaturen på vatten (Schijven 2001). 26 och organiskt material runt markpartiklarna. Löst organiskt och oorganiskt material sköljs bort. Här­ igenom minskar det ”bindningslager” som håller organismerna till partiklarna och en repulsion mellan de negativt laddade markpartiklarna och organism­ erna uppstår och underlättar en frisättning (Stenström 1985). 6.2.4 Jonkoncentration och jonsammansättning i vattnet Fasta ytor och material i den naturliga miljön är ofta negativt laddade. En adsorbents ytladdning är beroende av joniseringen av grupper på ytan eller adsorption av joner från lösningen runt materialet. Joniseringen av ytgrupperna bestäms av pH på lösningen (som tidigare diskuterats i 6.2.1 pH). Ad­ sorptionen av mikroorganismer blir bäst där störst skillnad i laddning råder mellan material och mikro­ organism och kan resultera i en irreversibel adsorpt­ ion om attraktionen är tillräckligt stark (Stevik et al. 2004). Om en hög koncentration av joner med en hög valens existerar i vattnet ökar i allmänhet reduktionen (Stenström 1985). En ökad katjonkoncentration neutraliserar partiklars och virus elektronegativitet och adsorptionen sker lättare. En sänkning av jonstyrkan kan ge en kraftig desorption (Stenström 1980). Detta styrks i Jörgensen m.fl. (2001), Guan m.fl. (2003), Stevik m.fl. (2004) samt Jin m.fl. (1997) vilka också hävdar att högre jonstyrka och valens, leder till ökad adsorption och minskad transport. Detta beror på att det ”diffusa elektriska dubbellagret”, Gouy-Chapmanlagret, mellan ytorna som skapar repulsion, blir svagare och tunnare vid hög jonkoncentration. Van der Waals krafter och andra attraherande krafter som finns på korta avstånd blir mer signifikanta och en fastläggning kan ske av organismen. Den kritiska koncentrationen över vilken jonstyrkan påverkar fastläggningen av mikroorgan­ ismer anses vara ca 10–100 mM för Na+ och ca 0,1–1 mM för Ca 2+. Om jonstyrkan i ingående vatten sänks, vilket kan ske efter kraftiga regn, snösmältning m.m., sker en viss frisättning av adsorberade organismer. Frisätt­ ningen beror på en minskad mängd positiva motjoner Figur 6-3. Schematisk bild av tillstånd vid ökad jon­ styrka. Zhuang och Jin (2003) undersökte fenomenet beskrivet ovan mer noggrant. De undersökte hur lösningens jonstyrka påverkade transporten av fagerna MS2 och φX-174 i kolonner med aluminiumoxidbelagd sand. Hög jonstyrka gynnade transporten i detta försök och resultatet kan förklaras med att ökad jonkoncentration skärmar av både laddningen på partikeln som ska adsorberas och adsorbantens laddning. Beroende på laddningen på partiklarna och ytan, blir effekterna av avskärmningen olika, se Figur 6-4. När partiklarna och adsorbantens yta har samma laddning, minskar en högre jonstyrka den elektrostatiska repulsionen mellan dem och adsorpt­ ionen ökar. P.g.a. samma avskärmande effekt kan ökad jonstyrka minska adsorptionen när partiklarna och adsorbanten har olika laddning eftersom avskärm­ ningen minskar den elektrostatiska attraktionen. I för­ söken var sanden positivt laddad p.g.a. beläggningen Figur 6-4. Jonstyrkans påverkan på adsorptionen av virus på en positivt laddad yta. 27 med aluminiumoxid gav sanden en pHiep på 9,0. Båda fagerna var negativt laddade då pH för lösningarna var 7,5. Ökad jonkoncentration minskade attraktionen mellan de olikladdade ytorna och trans­ porten genom kolonnerna underlättades. Inte bara jonkoncentrationen utan även samman­ sättningen av joner påverkar avskiljningen av virus. Flervärda katjoner kan binda virus och adsorbant med lika laddning genom att bilda saltbryggor mellan dem. Flera studier visar en ökad adsorption när fler­ värda katjoner finns närvarande i materialet till skillnad från envärda katjoner. Exponerade aluminium-, järn- och manganjoner ger högre adsorption. (Flynn et al. 2004). Finns det katjoner (Na+, Ca 2+, Mg2+) eller metalliska katjoner (Fe3+, Cu 2+, Zn 2+) i lösningen förhöjs adsorptionen av bakterier, medan an­ joner (Cl–, NO3–, SO42–) inte inverkar på adsorpt­ ionen (Stevik et al. 2004). Fosfat visade sig ha en negativ effekt på reduktionen av virus i kolonnförsök (Zhuang & Jin 2003). Fosfaten föreslogs ha skärmat av de positiva laddningarna på aluminiumoxid sanden och konkurrerat om bindningsplatserna. De tvåvärda jonerna Ca2+ och Mg2+ hade samma effekt, men med lite olika orsak; de hindrade attraktionen mellan de olika laddningarna på fagerna och sanden och minskade avskiljningen. på mikroorganismer är större än 5 % av diametern av partiklarna som utgör filtermaterialet (Foppen, Mporokoso & Schijven 2005). Om materialet är grövre och kanalbildning och makroporer är betyd­ ande minskar adsorptionen beroende på den kortare kontakt tiden och det ökade avståndet mellan materi­ alet och bakterierna (Stevik et al. 2004). Pang m.fl. (2005) refererar till en studie där silning/mekanisk filtrering endast var betydande då mikroorganism­ ernas storlek var större än 10 % av storleken på materi­ alet. Det sker mycket lite silning/mekanisk filtrering när mikroorganismens storlek är mindre än 8 % av materialets storlek. Alla partiklar som transporteras med vattnet och är större än 20 % av d10 blir kvarhållna menar Huisman och Olsthoorn 1983 (citerad i Fryklund 1992). Snabb reduktion har normalt erhållits i moränjordar enligt Stenström (1985). I siltig och sandig jord är överlevnaden längre. Detta beror förmodligen på skillnaden i pH i de olika jordarterna där morän överlag är mycket surare (Stenström 1985). Mikro­ organismerna överlever längre men adsorberas bättre i jordar med mindre partiklar som lera och silt, vilka har en större specifik ytarea och mer heterogen ladd­ ningsdistribution (Schjiven et al. 2001, s 31). Enligt DeBorde m.fl. (1999) är virus transporthastigheter beroende av materialet och materialets storlek. Hastigheterna kan variera mellan 1 m/d till 300 m/d beroende på om akvifärens material är permeabelt eller inte. 6.3 Egenskaper hos marken och infiltrationsmaterialet 6.3.2 Mineral- och Jordsammansättning 6.3.1 Partikel och porstorlek Nästan alla typer av sand består av flera mineraler, de är poly-minerala. Flynn, Rossi och Hunkeler (2004) redovisade skillnad i olika mineralers förmåga att reducera virus, genom att kolonner spikades med bakteriofagen H40/1. De mineraler som undersöktes var de som ingick i en sand tagen från en Schweizisk testanläggning för konstgjord grundvattenbildning. Det var en specifik fraktion av de olika mineralerna som analyserades för att kunna urskilja olikheterna hos mineralerna. De mineraler som testades var: kvartssand, granitsand, kalkstenssand (CaCO3) och kvarts-kalkstenssand. Det största genombrottet av H40/1 var för kvarts och granit, där upp till 98 % (91 % ±7 % resp. 92 % ±6 %) av tillsatta fager återfanns i uttaget vatten. Dessa resultat skiljde sig mycket från försöken för kalkstenssand som inte gav något Partikelstorleken hos materialet styr mycket av hur bakterier transporteras och relationen mellan storleken på patogenen och materialet är av betydelse. Silt, lera och fin sand har porstorlek som ligger i närheten av bakteriers storlek och leder därmed till silning/mekanisk filtrering. De små partiklarna (< 1 mm) är de som bidrar mest till adsorptionspro­ cessen, eftersom deras yta bidrar mycket till den totala ytan för ett sandprov och det är denna storleksfraktion som kan fokuseras på vid försök (Flynn, Rossi & Hunkeler 2004). Retentionen av bakterier som orsakas av filtrering är omvänt proportionell mot storleken på filter­ materialets kornstorlek. Generellt blir silning/mek­ anisk filtr­ering en betydande process när diametern 28 genombrott, dvs all H40/1 reducerades. Den mixade sanden (40 % kalkstenssand och 60 % kvartssand) gav ett litet genombrott på första spikningen (5 %) men avskiljningen försämrades vid de följande försöken. Försök gjordes även på sanden uppbyggd av olika mineraler under olika typ av behandling. Spikning med H40/1 i kolonner skedde på sand som dels var i ursprungligt skick och dels hade blivit tvättad med vatten. Den opåverkade sanden fick ett genombrott av fager på ca 40 %. Återanvändes denna, efter sköljning och torkning, kunde en tydlig försämring av reduceringsförmågan ses och genombrotten blev 40–80 %. Sanden som tvättades tre gånger innan spikning, fick även den en sämre förmåga att avskilja virus och ett genombrott på ca 85 %. Återanvänd­ ning eller tvättning tre gånger resulterade alltså i ca dubbelt så högt genombrott. Ett lager med mycket fint silt- och lermaterial på ytan av sanden tvättades bort vid de olika behandl­ ingarna. Detta resulterade i den sämre reduceringsförmågan. Sand med en fin beläggning adsorberade H40/1 bättre än de utan ett lager trots deras liknande mineralogi. Resultaten från försöken talar för att det fina materialet på kornytan reducerar de repellerande laddningarna på ytan tillräckligt mycket för att adsorption av den negativt laddade H40/l ska kunna ske. Borttagningen av beläggningen genom tvättning eller annan behandling resulterar i mindre ojämnheter/laddnings heterogenitet och gör adsorpt­ ionen av virus svårare (Flynn Rossi & Hunkeler 2004). Lerpartiklar är små med stor ytarea, ganska platta och har hög katjon-utbyteskapacitet vilket kan förklara varför fastläggningen blir stark av bakterier till lera. Även Thompson m.fl. (1998) påvisar denna tendens i batch-försök med tre olika sand- och jordblandningar. Adsorptionen av fagen φX-174 var störst till jorden med högst innehåll av organiskt material och lerpartiklar (9,21 mgOM/g, 9,5 % lera), näst högst till jorden med (6,7 mgOM/g, 4,5 % lera) och viss adsorption skedde till kvartssanden utan lerfraktion och organisktmaterial. ske på en liten del av partiklarnas yta, som har en laddning som främjar en fastläggning energimässigt. Dessa platser, som är ett resultat av heterogenitet i den totala laddningen, är små fläckar med positiv laddning. Flera studier talar för att virus föredrar ad­ sorption till delar av adsorbanten med gynnsamma laddningsegenskaper. Polariteten och styrkan på laddningen på en minerals yta (adsorbanten) är även den beroende av pH. Det pH där nettoladdningen är noll för mineralen utgör den isoelektriska punkten (Elimelech et al. citerad i Flynn Rossi & Hunkeler, 2004). Material med en hög isoelektrisk punkt är bättre adsorbanter än de med en lägre punkt (Schjiven 2001), eftersom de blir svagt negativt laddande eller positivt laddade vid naturligt pH och repulsionen blir mindre (Gerba & Bitton, citerad i Flynn Rossi & Hunkeler 2004). Ett exempel på detta är järn-, mangan- och aluminiumoxyhydroxider som har en hög pHiep och bildar positivt laddade områden på jordpartiklarna och erbjuder gynnsamma fastläggningsplatser för negativt laddade virus (Loveland et al. 1996 i Schjiven et al. 2000) Bättre reduktion vid liknande förhållanden har uppvisats vid kolonnstudier (Jin et al. 1997) och i fältstudier (Pieper et al. 1997 och Ryan et al. 1999 citerade i Schjiven et al. 2000). Efter regn finns tydliga indikationer på att virus desorberas, vilket kanske kan bero på förändring i laddningsförhållandena i marken, en ökad mängd organiskt material infiltreras och ett utbyte sker på sedimentens yta (Stenström 1980). Men det kan även bero på det låga innehållet av katjoner i regnvatten enligt Leclerc m.fl. (2000). 6.3.4 Hydrofobicitet Adsorptionen är styrd av hydrofobiciteten av sedimenten. Starkare hydrofobicitet leder till ökad adsorption av virus (Jörgensen et al. 2001). Chatto­ padhyay och Puls (1999) testade hydrofob adsorpt­ ion till olika mineraler och visade att bäst adsorpt­ ion var mellan den mest hydrofoba mineralen med den mest hydrofoba bakteriofagen. Är ett sediment hydrofobt innebär det att det finns fler ställen att binda andra hydrofoba partiklar och mikroorgan­ ismer och färre vattenmolekyler. Ju högre hydro­ fobicitet och jonstyrka och ju lägre pH i vattnet, desto bättre adsorption (Jörgensen et al. 2001). Detta vari­ erar dock stort med vilken typ av mikroorganism. 6.3.3 Laddning hos marken Naturligt infiltrationsmaterial i marken är till största del ogynnsamt för fastläggning av mikroorganismer, p.g.a. den tidigare beskrivna negativa laddningen och andra egenskaper. Adsorptionen tros därför endast 29 Växelverkan mellan hydrofoba grupper på ytan på mikroorganismen och adsorbanten kan främja adsorption. Vid höga pH, när båda partiklarnas ytor är negativt laddade kan det vara hydrofob interaktion som är orsaken till fastläggningen (Schjiven 2001). Men Chattopadhyay och Puls (1999) hävdar att även vid pH mycket nära pHiep för både partiklarna som ska adsorberas och adsorbenten är hydrofobiska inter­ aktioner det som styr adsorptionen. Se även avsnitt 6.1.5. 6.4.1 Uppehållstid Uppehållstiden för grundvattnet i marken har en viktig påverkan på reduktionen av patogener. I en studie om inducerad infiltration där uppehållstiden var två resp. tio veckor vid två olika anläggningar med i övrigt liknande förutsättningar var avskiljningen överlag två tiopotenser bättre för den längre uppe­ hållstiden (Havelaar, van Olphen & Schijven 1995). I de anläggningar som studerades av Sundlöf och Kronkvist (1992) visade sig uppehållstiden ha betyd­ else för reningsgraden av organiskt material, och skulle därigenom även kunna påverka reduceringen av mikroorganismer. Anläggningar med uppehållstid över 50 dagar hade bäst avskiljning av det organ­ iska materialet. Uppehållstid är ett mått som innefattar och påverkas av avståndet på infiltrationen, material och permeabilitet, omättad zon och ytbelast­ ning hos anläggningen. Materialet kan påverka uppe­ hållstiden, men det är främst hur infiltrationen sker och hur anläggningen drivs som kan vara begräns­ ande. Vissa avvikelser från detta samband kunde i denna studie förklaras med att uppehållstiden endast hade skattats. Halterna som reningsgraden beräknades från, redan innan var mycket låga och extrema förhållanden för andra parametrar påverkar resultatet från några av anläggningarna. 6.4 Anläggningens utformning och drift De faktorer som diskuteras i detta kapitel, uppehållstid, transportsträcka, grundvattenhastighet och vattenmättnadsgrad, är även analyserade i kapitel 7 Kvantifiering av de dimensionerande faktorerna och är endast översiktligt och kvalitativt beskrivna här. En kort genomgång av biohudens och naturliga mikro­organismers betydelse görs. Figur 6-5. Avskiljning av MS2 och φX-174 i relation till uppehållstid i olika akvifärer (Schjiven 2001, s.207). Data är tagen från flera olika fältstudier. 30 Figur 6-5 visar ett exempel på fagers avskiljning i olika akvifärer. Reduktionen sker fortast i början och avtar därefter. Det skiljer sig mellan en och tio dagar innan en 6 log10 reduktion har skett för fagerna i de olika akvifärerna. Detta beror på att förhållandena och egenskaperna (pH, partikelstorlek, porhastighet, hydraulisk konduktivitet, joninnehåll osv.) hos akvi­ färerna är olika. hur avskiljningen av virus avtar med avståndet utifrån flera studier (se figur 6-6 nedan). Gerba och Lance (refererade i Schjiven 2001, s. 62) uppmätte under ett kolonnförsök en avskiljning på 1 log10 (C/C0 = 0,1) de första 5 cm, men det krävdes 35 cm för att få ytterligare en log10. Under ett fullskaleförsök utfört av Schjiven (2001, s.76) avskiljdes ca 3 log10 de första 2,4 m och sedan 5 log10 linjärt under de följande 27 m. Även Deborde m.fl. (1999) fick liknande resultat vid ett fullskaleförsök, men det skiljer mycket mellan olika anläggningar och akvifärer, vilket diskuteras i kapitel 7. Det icke-linjära förhållandet mellan reduktion och transportsträcka kan leda till att vissa studier i pilot­ skala överskattar reduceringsförmågan för en sand under givna betingelser. Uppskattningar av virusreduktion för längre avstånd utifrån kolonnstudier eller från försök i fält med kort avstånd kan bli grovt överskattade (Schijven et al. 2000). Det längsta avståndet som en mikroorganism eller virus kan transporteras beror som ovan på grundvattnets hastighet, överlevnad, koncentration i infiltrerande vatten, utspädning och dispersion. Gerba m.fl. (1984 citerad i Jin et al. 1997) fann virus ca 30 meter ner i marken och på 100 m avstånd från infiltration av avloppsvatten. Vaughn och Landry (1977 citerad i Jin et al. 1997) fann coxsackievirus ca 400 m från en deponi med avloppsslam. Gerba m.fl. (1984 citerad i Stevik 2004) fann att koliforma bakterier kunde transporteras mellan 1–450 m i olika jordar. Enligt Noonan och McNabb (1979) kan virus transporteras över 900 m i en akvifär med grovkorn­ igt material (citerad i DeBorde 1999). Ett av de längsta avstånden som modell organismer för virus 6.4.2 Transportsträcka mellan infiltration och uttag Patogener, andra mikroorganismer och ämnen reduceras i kontakt med materialet vid konstgjord grundvattenbildning. Avståndet från infiltrationen till uttagspunkten blir därmed en avgörande parameter för hur mycket som avskiljs. Eftersom det är så många faktorer som inverkar är det en stor skillnad mellan olika akvifärer och anläggningar, vilket påverkar transportsträckan som krävs för att få fullgod reduktion. Därför kunde Sundlöf och Kronkvist (1992) inte visa korrelation mellan reningsgrad av organiskt material och avståndet på infiltrationen för de studerade anläggningarna, med den begränsade infiltration som fanns tillhanda. Effektiviteten av filtreringen tenderar att avta med avstånd och tid från källan. Avlagringar av suspend­ erade partiklar och filtrerat material orsakar igensättningar och ett lager bildas redan den första metern som försvårar mikrobiologisk transport. Reduktionen av mikroorganismer är mest betydande de första metrarna och avtar med avståndet (Pang et al. 2003; Schjiven et al. 2000). Schjiven (2001. s.64) beskriver Figur 6-6. Kollisionseffektivitet, α, mot sträckan. Kollisionseffektiviteten är andelen mikroorganismer eller partiklar som kolliderar med adsorptionsmaterialet och fastnar. (Schjiven 2001, s. 63). 31 Ett ökat porvattenflöde komprimerar det hydrodyn­ amiska gränsskiktet runt partiklarna vilket kan ge en bättre sorption av mikroorganismerna till part­ iklarna. Detta betyder att vid högre belastningar fastnar mikroorganismerna bättre i små porer, dock blir den sämre i de större pga mindre kontakttid med sidorna (Bengtsson 2005). upptäckts från källan är 1250 m. Auckenthaler, Raso och Huggenberger (2002), fann fagerna H4 och H40 på detta avstånd vid transport i en kalkstensakvifär med en grundvattenhastighet på 800 m/dag. 6.4.3 Grundvattenhastighet Hastigheten på det infiltrerade vattnet har betydelse för reduktionen av patogener. Om hastigheten på grundvattnet är hög ökar genomströmningen av vatten i de större porerna och kontakttiden mellan mikroorganismerna och materialet blir mindre och filtreringen och adsorptionen av mikroorganismer minskar (Stevik et al. 2004; Schijven 2001, s. 222). Porhastighetens inverkan på reduktionen studerades av Diezer m.fl. (2005) i en pilotinfiltrationsanläggning. Bestämning av genombrottshastigheter och tot­ alt genombrott gjordes, definierat som genombrott efter fem dygn. Porhastighet är en viktig parameter för reduktionen av mikroorganismer och är omvänt proportionell. Ökades hastigheten fyra gånger, från 180 till 720 cm/d, steg det totala genombrottet med 350 ggr för indikatororganismerna somatiska fager och över 1000 ggr för F+fager och E.coli. I Schjivens kolonnförsök ökades porhastigheten till det dubbla (från 1,7 till 3,3 m/dag) och det resulterade i en avskiljning som var 2 log10 lägre. Kolonnförsök som genomfördes av Matthess m.fl. (1991 citerad i Foppen, Mporokoso & Schijven 2005) visade på två olika typer av genombrott av E.coli vid olika porvattenflödeshastigheter. Vid låga flöden, 0,7–1,5 m/dag, blev genombrottet litet och planade ut i en platå. Vid högre flöden, 2–7,5 m/dag, var genombrottet snabbare och C/C0 värden högre. Men enligt Pang m.fl. (2003) resulterar en ökad porhastighet, om ett homogent material antas och inga makroporer förekommer, i en ökad första ordningens filtrationshastighet. Pang m.fl. (2003) visar i sina pilot­ försök att avskiljningshastigheten per dag ökade med en högre porhastighet. Enligt ”colloid filtration theory” är avskiljningen (reduktionsfaktorn, definierad som log10 reduktion/m) proportionell mot v–2/3 (v i m/dag) och avskiljningen per tid, (log10 re­duktion/dag) pro­ portionell mot v1/3 (Yao et al. 1971 i Schjiven 2001, s. 151). Reduktionen per meter minskar alltså enligt denna teori med en ökad hastig­het men reduktionen per dag skulle alltså öka. Så förklarar Pang m.fl. (2003) sina resultat, som under sitt försök antog att det var mekanisk filtrering som styrde reduktionen. 6.4.4 Vattenmättnadsgrad Flera undersökningar tyder på att omättat flöde i en anläggning för konstgjord grundvattenbildning är att föredra vid infiltration och att den omättade zonen stärker beredningseffektiviteten (Stevik et al. 2004; Jörgensen et al. 2001; Leclerc et al. 2000; Yin et al. 2000; Chu et al. 2003; WHO 2004). Om ett mättat flöde uppstår vid infiltrationen blir reningen av vattnet sämre och omvandlingen av ytvatten till grundvatten sker inte till lika stor del. Det mättade flödet påverkar transporten så att den till största del sker i de grova porsystemen och kontakten mellan markpartiklarna och patogenerna blir sämre och adsorptionen försämras. Om genomströmningshastigheten ökar, minskar uppehållstiden och tiden för den mikrobiella nedbrytningen (Hanson 2000). Vattenmättnadsgraden i en mark påverkar redukt­ ionen av patogener både genom att påverka silning och filtrering samt genom att påverka mikroorgan­ ismernas inaktivering och avdödning. Avdödningen av mikroorganismer och inaktivering av virus har visat sig vara försumbar, jämfört med reduktion genom adsorption och filtrering, vid pilotförsök med infiltration i mättade kolonner. Under omättade för­ hållanden är bidraget av avdödningen till den totala reduktionen större (Jin, Chu & Li 2000). Enligt Sundlöf och Kronqvist (1992) finns en god korrelation mellan reningsgrad av organiskt material och omättad zon upp till en viss begränsad mäktighet. Undersökningen av tjugo anläggningar visar att ett samband finns för de anläggningar som har en mäktighet av den omättade zonen mellan 0 till 5–6 m. Inkluderas de anläggningar som har en omättad zon som överstiger 6 meter är sambandet inte lika tydligt. En kort omättad zon kan kompenseras med låg ytbelastning visar studien. Makroporer eller kanalbildning i infiltrationsmaterialet minskar filtrationen och har visat sig påverka transporten av bakterier. Vid omättad zon sker den mesta transporten i de mindre porerna och 32 betydelsen av makroporerna blir mindre (Stevik et al. 2004). Transport av bakterier sker endast i vattenfyllda porer enligt Jörgensen m.fl. (2001). Därför är kopplingen mellan porerna och vatteninnehållet i marken viktigt. Ju längre sträcka en organism måste färdas för att nå ner till grundvattnet, desto mindre risk att den verkligen når fram. I den omättade zonen blir denna sträcka längre än i den mättade menar Hanson (2000). Fenomenet med ökad reduktion i ett omättat syst­ em har tillskrivits närvaron av ett gränsskikt mellan luft och vatten (Air-Water Interface, AWI) som inte finns vid mättade tillstånd (Jin & Chu 2000; Schijven 2001 s. 53, Schäfer et al. 1998). Det finns inga direkta bevis, men försök tyder på att den ökade reduktionen av virus vid omättade förhållanden kan bero på inaktivering vid gränsskiktet. Gränsskiktet påverkar virus så att dess yttre membran av proteiner ändras vilket leder till att viruset blir mindre benäget att infektera (Jin et al. 2000). Detta sker genom att hydrofoba delar av viruset sträcker sig över till gasfasen vid gränsskiktet genom att förändra sin kapsid­ struktur. Förändringen eller rekonfigurationen resulterar i minskad infektivitet hos viruset (Thompson et al. 1998). Gränsskiktet verkar vara anledningen till den ökade reduktionen av MS2 och X174 vid omättade förhållanden och speciellt inaktiveringen av MS2. Till vilken utsträckning ett virus fastnar på skicktet är också beroende av den isoelektriska punkten för ett virus, men resultaten tyder på att adsorption lättare sker till materialet vid omättade förhållanden (Jin & Chu 2000). Det finns en variation av tåligheten hos olika mikroorganismer för både högt och lågt vatteninne­ håll, men generellt är överlevnaden längre i fuktig mark än i torr. De längsta överlevnadstiderna för Streptococcus faecalis, var under mättade förhåll­ anden (Kibbey et al. citerad i Stevik et al. 2004). Försök visar att inaktivering av virus når ett maxima nära markens fältkapacitet. Tillsats av organiskt material med infiltrationsvattnet kan bidra till att inaktiveringen försämras. Det organiska materialet fastnar och mättar gränsskiktet mellan luften och vattnet, vilket hindrar virus från att fastna och inaktiveras i gränsskiktet (Schjiven 2001, s. 53). Över­ levnaden för mikroorganismer skiljer sig om de är fria i ett vatten eller adsorberade på en yta. Pang m.fl. (2003) visar genom pilotförsök att avdödningen var 4–6 ggr högre för adsorberade F-RNA fager och 15–28 ggr högre för E.coli bakterier. 6.4.5 Biohud och naturliga mikroorganismer De är flera studier som visar att det är i det översta lagret i marken som den största delen av reduktionen sker och tyder på en stor påverkan av naturliga mikro­ organismer, som i huvudsak förekommer i de översta marklagren (Jensen et al. 2005). Uppbyggnaden av en biofilm minskar porstorleken och en filtrering blir mer effektiv. Det finns tydliga bevis på att avskiljningen av bakterier är bättre i system med en välutvecklad biofilm och mindre igensättningar än system utan biofilm (Stevik et al. 2004). De naturliga mikroorganismerna och nedbrytarna av det organiska materialet inverkar på patogenerna och reducerar deras överlevnadstid avsevärt. Den initiala avdödningen är högre och den totala tiden under vilken man kan detektera organismerna kort­ are om naturliga mikroorganismer finns närvarande (Stenström 1985). De naturliga mikroorganismerna verkar vara den mest betydande faktorn på virus överlevnad i grundvatten jämfört med temperatur, syra och näringsinnehåll enligt Gordon och Toze (2003 citerad i Rzeżutka & Cook 2004). Den natur­ liga mikrofloran påverkar nedbrytningen av bakterier och virus och det har visat sig i försök att en bättre nedbrytning sker med en bred population av ursprungliga bakterier i grundvattnet än om mikroorganismerna testas individuellt (Wall et al. 2005). 7 Kvantifiering av de dimensionerande faktorerna 7.1 Inledning Ett syfte med litteraturstudien var att försöka kvanti­ fiera den mikrobiologiska barriärverkan vid olika förutsättningar vid konstgjord grundvattenbildning utifrån resultat från tidigare undersökningar som gjorts i fullskaliga anläggningar och försöksområden. Kvantifieringen ska förhoppningsvis kunna bidra i 33 bedömningen av befintliga anläggningars mikrobio­ logiska barriärverkan och dimensionering av framtida anläggningar. 7.3 Reduktion i förhållande till transportsträckan Reduktion med avseende på avståndet mellan in­ filtrationsplats och uttag varierar mellan akvifärer beroende på olika förhållanden och egenskaper, se Figur 7-1. Det är stor variation mellan akvifärerna; vid 20 m ligger intervallet mellan ca 3–8 log10 reduktion och för 40 m mellan ca 4–8 log10. De lång­ samma akvifärerna hamnar i den övre delen av spannet efter redan 20–30 m; det korta avståndet kompenseras med en längre uppehållstid och en lång transport blir inte nödvändig. De ”snabba” akvifär­ erna, Burnham och Erskine, behöver ett längre avstånd för att komma upp i en reduktion runt 5–6 log10. De höga värden på reduktion efter ett kort avstånd som fås från Someren jämfört med de andra studierna kan bero på att processen är s.k. djupinfiltration, vilket medför att betingelserna är lite annorlunda jämfört med de andra studierna. Där­ emot hamnar både R5 från Somerenstudien och enterovirus i Remmerdenstudien utanför ”spannet” för långsamma akvifärer, vilket beror på att de är svåra att avskilja och mycket persistenta. 7.2 Metod För att försöka göra en uppskattning av reduktionen har data samlats från fältstudier från olika länder som utförts under naturliga förhållanden. Spikning har skett med bakteriofager av olika typer och ibland även bakteriella indikatororganismer. Urvalet av fält­ studier var tämligen enkelt, de som analyserats är de som redovisats som artiklar och inte haft except­ ionella förhållanden, och det är inte många studier som ut­förts på detta sätt genom spikning i fullskaliga akvifärer eller riktiga anläggningar. Referenser finns angivna i Bilaga B. De studerade anläggningarna är ganska skilda i utformning och karaktäristik. Fyra av studierna är utförda vid bassänginfiltration, ett vid inducerad infiltration och ett vid djupinfiltration. De olika förutsättningarna på anläggningarna avspeglas på resultaten som är mycket varierande. Tre av studierna är utförda i Nederländerna, vid anläggningarna Castricum (bassänginfiltration), Remmerden (inducerad infiltration) och Someren (djupinfiltration). De är tre akvifärer med en grundvattenhastighet på ca 1–3 m/dag, som har fått beteckningen ”långsamma” i denna studie. Materialet är fin sand (0,2– 0,27 mm) med stråk av lera och silt (ca 1 % i de olika akvifärerna). Kokemäenjoki-studien i Finland är gjord i en långsam artificiell akvifär (avgränsad med plast), efter att vattnet infiltrerades i en omättad gruskolonn med hög genomströmning. De två andra studierna är utförda i Erskine, Missoula USA, och Burnham, Nya Zeeland och i dessa akvifärer består materialet av grövre stenkross och grus (upp till 15 cm) och har en hastighet som överstiger 25 m/dag och klassas i denna litteraturstudie som ”snabba” akvifärer. Reduktionen har analyserats med avseende på av­ ståndet till uttagsbrunnarna och uppehållstiden, efter­ som avstånd i två olika akvifärer kan innebära stora skillnader i uppehållstid. Beräkningar har även gjorts av en s.k. reduktionsfaktor, vilken reduktion som sker per meter vid olika avstånd från injektionsbrunnen. 7.4 Reduktion i förhållande till uppehållstid Även för uppehållstiden är variationen stor för hur mycket som reduceras i de olika anläggningarna, se Figur 7-2 och 7-3. Efter två dagar är reduktionen mellan 3–8 log10. Efter 14 dagar, vilket är gränsen för att klassificera ytvatten som grundvatten i Sverige, ligger reduktionen mellan 4–8 log10. De snabba akvifärerna, Burnham och Missoula (markerade med fyrkanter respektive cirklar i diagrammet), kommer upp i en ganska hög, ca 5–8 log10 redan efter ett och ett halvt dygn, beroende på vilken organism som studeras. Studien visar att detta antagligen beror på det långa avståndet som transporten sker, vilket kompenserar den korta uppe­ hållstiden och ger en tillräcklig kontakt med materi­ alet. 34 9 "Långsamma", 1-3 m/dag 8 "Långsamma" 7 6 5 4 3 "Snabba" Reduktion (-log(C/C0)) Castricum - MS2 Remmerden - FRNA Remmerden - Enterovirus Someren - R5 Someren - PRD1 Someren - MS2 Someren - E.coli Kokemäenjoki - MS2 Burnham - MS2 Burnham - B. subtilis Burnham - B.subtilis 2 Erskine - MS2 Erskine - X174 Erskine - Polio Erskine - PRD1 "Snabba", >25 m/dag 2 1 0 0 10 20 30 40 50 60 70 80 90 100 Avstånd (m) Figur 7-1. Reduktion mot transportsträcka. De snabba akvifärerna är punkter och fyrkanter, de långsamma är trianglar och sneda fyrkanter. 9 8 Castricum - MS2 Remmerden - FRNA Remmerden -Enterovirus Someren - MS2 Someren - PRD1 Someren - E.coli Someren - R5 Kokemäenjoki - MS2 Burnham - MS2 Burnham - B.subtilis Burnham - B.subtilis 2 Erskine - MS2 Erskine - Polio Erskine - X174 Erskine - PRD1 "Långsamma" 6 5 4 3 "Snabba" Reduktion (-log(C/C0)) 7 2 1 14 dygn 0 0 5 10 15 20 25 30 35 40 Uppehållstid (dygn) Figur 7-2. Reduktionen i olika fältstudier i förhållande till uppehållstiden, 14 dagar är markerat med ett vertikalt streck. Den sneda linjen representerar en uppskattad lägsta avskiljning, dvs. ”worst case”. 35 9 8 Castricum - MS2 Remmerden - FRNA Remmerden -Enterovirus Someren - MS2 Someren - PRD1 Someren - E.coli Someren - R5 Kokemäenjoki - MS2 Burnham - MS2 Burnham - B.subtilis Burnham - B.subtilis 2 Erskine - MS2 Erskine - Polio Erskine - X174 Erskine - PRD1 6 "Långsamma" Reduktion (-log(C/C0)) 7 5 4 3 "Snabba" 2 1 0 0 2 4 6 8 10 12 14 16 Uppehållstid (dygn) Figur 7-3. Reduktion i förhållande till uppehållstid, figur 7-3 med skalan <16 d. Den sneda linjen markerar skill­ naden mellan ”långsamma” och ”snabba” akvifärer. 1,4 Castricum - MS2 Remmerden - FRNA 1,2 Remmerden - Enterovirus Reduktionsfaktor (-log(C/C0))/m Someren - MS2 1 Someren - PRD1 Kokemäenjoki - MS2 Burnham - MS2 0,8 Burnham 2 - B.subtilis Burnham - B.subtilis Erskine - PRD1 0,6 Erskine - X174 Erskine - Polio 0,4 Erskine - MS2 0,2 0 0 10 20 30 40 50 60 70 80 90 100 Avstånd (m) Figur 7-4. Reduktionsfaktor vid olika avstånd. 5 m och 30 m markerade i bilden. studierna, se Figur 7-4. Det blir tydligt att redukt­ ionen inte är linjär, vilket är viktigt att ta hänsyn till vid dimensionering av anläggningar. Efter ca 5 meter är reduktionen mellan ca 0,3–0,9 log/m och redukt­ ionsfaktorn minskas till ca 0,1–0,3 log/m efter 30 m. Beräkningen har gjorts genom att aktuell reduktion vid ett visst avstånd delas med hela transportsträckan, vilket blir ett medel av reduktionsfaktorn upp till det avståndet. För dimensionering innebär detta ett 7.5 Reduktionsfaktor Liksom för faktorerna ovan är det mycket stora skill­nader i vilken reduktion som sker per meter vid olika avstånd (s.k. reduktionsfaktor) mellan de olika Reduktionsfaktor; (–log10(C/Co)/m), filter factor på eng. även removal rate vid modellering. 36 avstånd på mellan ca 25 och 70 m för en reduktion på 7 log10 beroende på om det är en snabb eller lång­ sam akvifär, om ett antagande görs att reduktionen efter 30 m kan användas som ett genomsnitt. 7.7 Diskussion av kvantifierings­ studien och jämförelse med lagstiftning internationellt Utifrån kvantifieringsstudien kan det tyckas som om det inte finns en rimlig grund för att bestämma säkerhetsavstånd eller uppehållstid för infiltrationen vid vilken en säkerhetsbarriär kan garanteras. Genom­ gången av de olika studierna visar att det finns mycket osäkerhet i bedömning av den mikrobiologiska barri­ ären vid ett vattenverk med konstgjord grundvatten­ bildning. Dock finns det trender i resultaten som kan skapa förutsättningar för en bedömning. För att dimensionera en anläggning optimalt utifrån ett perspektiv om tillräcklig reduktion krävs det ingående kunskap om området och akvifärens förhåll­ anden och förutsättningar. Stenström (1980) hävdade att det var svårt att dra några generella slutsatser och att komplexiteten i samspelet mellan alla olika faktorer som påverkar gör det mycket svårt att upprätta enhetliga regler för säkerhetsavstånd och riskbedömningar. Enligt Jörgensen m.fl. (2001) finns det ingen vetenskaplig grund för att sätta en specifik minimal uppehållstid för att garantera ett hälsomässigt säkert dricksvatten. De länder som har en utbredd användning och forskning om konstgjord grundvattenbildning har dock en viss lagstiftning och riktlinjer för anläggningar och säkerhetsbarriären i processen utöver generella dricksvattenföreskrifter, men dessa är lika varierande som studierna ovan. I Sverige gäller att råvatten, som härrör från konstgjord infiltration av ytvatten, normalt kan betraktas som opåverkat grundvatten om vattnets verkliga medeluppehållstid mellan infiltrations- och uttagspunkterna är 14 dagar eller längre. Om det är tveksamt om ett råvatten är ytvattenpåverkat eller inte, bör kemiska, fysikaliska eller biologiska metoder användas för att närmare ut­reda saken. Det finns dock inga allmänt accepterade metoder eller kriterier för att avgöra om ett råvatten är ytvattenpåverkat (Livsmedelsverket 2004). Vidare beror antal barriärer som krävs vid dricksvattenberedning på vilken typ av råvatten som används. Till exempel krävs en barriär för opåverkat grundvatten och för ytvattenpåverkat grundvatten, två eller tre för ytpåverkat grundvatten och för 7.6 Modellorganismer I studierna är modellorganismerna utvalda eftersom de är kända som persistenta och för att de transport­ eras lätt genom marken. Bakteriofagerna diskuteras i avsnitt 3.3.1. B.subtilis sporer är i samma storleksordning som bl.a E.coli, enterokocker, och salmon­ ella men är kända för att vara tåliga i naturliga miljöer. Den reduktion som kunde konstateras i Burnham studien ansågs därför ske endast genom silning och filtrering (Pang et al. 2005). R5, sporer av Cl. bi­ fermentans, används som modellorganism för oocyst­ er av Cryptosporidium parvum. Även om R5 är ca fem ggr mindre (ca 1 μm i diameter) än protozoer, är den lika tålig i vattenmiljöer och dess mindre storlek gör att den kommer att kollidera mindre med materialet och transporteras lättare och därmed är den en konservativ modellorganism (Schjiven et al. 2000). Enterovirus som mäts i Remmerdenstudien består till största del av Coxsackie B virus och ex­ isterar med de naturligt förekommande halterna. Det poliovirus som använts i Erskineförsöket är en försvagad typ. En av bakterierna, R5, som finns med i kvantifieringsstudien är sporbildande, vilket gör den jämförbar med tåliga parasiterna. Försöket gjordes i grovt material, vilket kan tolkas som att detta försök är ett ”värsta-scenario”. För de olika studierna är det ganska svårt att dra några slutsatser om de olika typerna av organismer; organismerna är mycket lika varandra och endast ett fåtal olika arter är använda. Bakteriofagerna som agerar modellorganismer ska helst ligga under avskiljningen av patogener, för att agera som konserv­ ativa modellorganismer. Modellorganismerna kan även jämföras med E.coli i studien Someren. E.coli är en indikatororganism som inte är lämplig för den här typen av undersökning. Eftersom E.coli inte är lika persistent och resistent som andra patogena organismer som kan transportera sig genom mark, är de fullständigt reducerade redan efter 8 m. Råvatten syftar här på det vatten som ska användas till att bereda dricksvatten och inte det ytvatten som ska infiltreras. 37 ytvatten beroende på bakteriekoncentrationen i rå­ vattnet. I Australien, Tyskland och Nederländerna används 50–60 dagars minimum uppehållstid som ett riktvärde för anläggningar (Jörgensen et al. 2001). Finland planerar en uppehållstid på 3 månader för nya anläggningar som ska producera dricksvatten (Niemi et al. 2004). Nya Zeeland har enligt Pang m.fl. (2003) godtyckligt uppskattat erforderligt avstånd mellan sept­ iska system och ytvatten till 20–30 m och mellan septiska system och grundvatten till 30–50 m, vilket kan tyckas kort eftersom bakteriekoncentrationen är hög om läckage skulle uppstå. Genomgången av litteratur tyder dock på att reduktionen kan styras med hjälp av faktorerna uppe­ hållstid, transportsträcka och grundvattenhastighet, vilket gör att gränsen på 14 dagar som är satt för att klassificera infiltrerat ytvatten som opåverkat grundvatten vid infiltration i Sverige inte är absolut. Reduktionen kan vid denna tidpunkt både vara till­ räcklig eller inte. Varken avstånd eller uppehållstid räcker för att dimensionera en anläggning var för sig. Studien pekar på att avståndet är en lika kon­ sekvent parameter, där ett avstånd mer än 40 m ger en reduktion på 4–8 log10 oberoende av grundvattnets hastighet. Livsmedelverkets rekommendation om 14 dagars uppehållstid skulle således kunna komplett­ eras med en rekommendation om ett minimalt avstånd mellan infiltrationspunkt och uttag på 40 m. Jämförs detta med beräknad önskad reduktion av virus enligt resonemanget om en acceptabel risk på en smittad person av 10000 per år (se avsnitt 3.2), kan det tyckas lite lågt, eftersom en önskad reduktion var 5–8 log10 för virus i ett råvatten i Nederländerna. Men det är samstämmigt med det intervall av reduktion som har skett efter 14 dagar, vilket också är 4–8 log10. Eftersom en mikrobiologisk säkerhetsbarriär ska ”motverka förekomst av sjukdomsframkallande virus, bakterier, och parasitära protozoer” (Citat från Livs­ medelverket 2004), inkluderas kort konstgjord infiltration av ytvatten (kortare tid än 14 dagar) som ett beredningssteg och säkerhetsbarriär vid dricksvattenberedning. Barriärverkan för en kort infiltration kan för t.ex. fem dagar uppskattas till > 3 log10 enligt fig. 7-3. Om detta jämförs med andra beredningsmetoder som räknas som barriär enligt Livs­ medelsverket (2004) ligger avskiljningen för kort konstgjord infiltration inom samma intervall som för kemisk fällning men är lite sämre än membranfiltrering. 7.8 Begränsningar Det är ganska få studier som är redovisade, med mer material skulle kanske ett ännu tydligare spann kunna urskiljas för en akvifär med ett visst material eller grundvattenhastighet. Några studier med en grundvattenhastighet som ligger mellan de hastigheter som har betraktats här har inte funnits i stud­ erad litteratur. Ingen kvantifiering av reduktionen som sker i den omättade zonen har kunnat genomföras eller studeras i litteraturen, vilket är en stor brist om den är så pass betydande som många studier tyder på. Det är en ytterliggare faktor som direkt påverkar utformningen och bedömningen av en anläggning förutom uppehållstid, transportsträcka, hastighet. För ett bidrag till resonemang om den omättade zonen vid konstgjord grundvattenbildning se kommande VAForsk rapport Reduktion av naturligt organiskt mater­ ial och mikroorganismer i konstgjord grundvatten­ bildning10. En liknande genomgång var tänkt att genomföras utifrån försök gjorda i kolonner/pilotstudier. Result­ aten blev mycket varierande och framför allt beräkningen av reduktionsfaktorn som var mellan 0,085 och 37 log10 reduktion/m för de olika försöken. Detta visar på osäkerheten i denna typ av försök och svårigheten att överföra kunskapen till verkliga förhållanden. De mest verklighetstrogna, naturliga förhållanden hamnade mellan 0,01–1 log10 redukt­ ion/m, vilket stämmer bra överens med fältstudierna. 1–10 st E.coli/100 ml och 10–100 st koliforma bakterier/ 100 ml kräver två barriärer, >10 st E.coli/100 ml och >100 st koliforma bakterier/100 ml kräver tre barriärer (Livsmedelsverket 2004). 10 Måns Lundh, Emma Holmström, Jonas Långmark, Henrik Rydberg, under arbete 2005. 38 8 Sammanfattning 8.1 Påverkande faktorer För att sammanfatta de olika faktorernas påverkan på den mikrobiologiska barriärverkan har tabell 8-1 sammanställts. Tabell 8-1 Faktorer som påverkar den mikrobiologiska barriären. Egenskaper hos mikroorganismen Koncentration −Generellt ökar silning och filtrering med högre koncentration av mikroorganismer, tills materialet blir mättat. Organismtyp −Det är stor variation på reduktionen mellan olika typer av mikroorganismer, vilket bl.a. beror på faktorerna redovisade nedan. Storlek −Storleken på mikroorganismerna påverkar silningen; större mikroorganismer har färre porer att ta sig igenom, och adsorptionen; större ytarea ger fler chanser till interaktion med materialet. Laddning hos mikroorganismen −Bestämmer den elektrostatiska interaktionen med adsorbenten och styrs av pH. Mikroorganismer och material är ofta negativt laddade i naturliga miljöer och repellerar varandra. Hydrofobicitet −Påverkar de hydrofoba interaktionerna mellan mikroorganism och material, kan vara starkare än de elektrostatiska interaktionerna. Egenskaper hos vattnet pH −Generellt ökar filtrering vid minskat pH, men detta beror på egenskaper hos mikroorganismerna och materialet. Överlevnaden av mikroorganismerna påverkas endast vid höga (>10) och låga (<5) pH. Temperatur −Vid lägre temperaturer överlever mikroorganismerna och virus längre, men filtreringen blir bättre. Innehåll av organiskt material −En hög koncentration av organiskt material minskar adsorptionen av mikroorganismer pga. att det konkurrerar om hydrofoba adsorptionsplatser. Jonkoncentration och jonsamman­ sättning −Högre jonstyrka och valens leder i allmänhet till ökad avskiljning. Egenskaper hos marken Partikelstorlek och porstorlek −Fint material ökar silning och filtrering, pga. de mindre porerna och större ytarea för adsorption. Inaktiveringen minskar dock. Mineral och jordsammansättning −Kvarts och granit adsorberar virus sämre än kalksten. Mineraler med ett fint ytskikt ökar adsorptionen. Laddning hos marken −Laddningen är beroende av pH, och material med hög pHiep är generellt bättre adsorbenter. Hydrofobicitet −Starkare hydrofobicitet hos materialet leder till ökad adsorption av mikroorganismer. 39 Forts. Tabell 8-1. Konstruktion och drift Uppehållstid − Reduktionen ökar med längre uppehållstid. Transportsträcka − Reduktionen ökar med längre transportsträcka. Grundvattenhastighet −Reduktionen under ett visst avstånd minskar med ökad grundvattenhastighet. Vattenmättnadsgrad − Silning och filtrering ökar vid omättade förhållanden. Biohud och andra mikroorganismer −En välutvecklad biohud främjar reduktionen, naturliga mikro­ organismer påverkar inaktivring och avdödning av patogenerna. Flera av parametrarna hänger ihop, t.ex. laddning och pH. Det är svårt att visa på vilken faktor som är mest avgörande för reningen. En slutsats som kan dras är att laddningarna på mikroorganismerna och virus som ska adsorberas, laddningen i lösningen och laddningen på markmaterialet (adsorbanten) påverkar avskiljningen. Eftersom laddningen på en yta styrs mycket av pH, blir denna faktor kritisk. Om akvifären är ”långsam” och endast ett litet uttag av vatten behövs, kan dimensionering ske enligt de vänstra spalterna i tabell 8-2, vilken har samman­ ställts med hjälp av figur 7-1, 7-2 och 7-3. Liknande resonemang har förts för en snabb akvifär. Studien föreslår att avstånd verkar vara en tillförlitlig parameter för att styra reduktionen. Enligt tabell 8-2, och de figurer den bygger på, tycks ett avstånd på över 40 meter vara tillräcklig för en reduktion på mellan 4–8 log10 av virus, vilket ger uppehållstider på mellan 1,5–35 dygn beroende på akvifärens karaktär. Möjligen skulle det därför kunna vara en idé att komplettera 14-dagars rekommendat­ ion från Livsmedelsverket med ett rekommenderat avstånd mellan infiltrations- och uttagspunkter. Denna rekommendation skulle kunna hamna på ca 40 m. 8.2 Kvantifieringsstudien Reduktionen av mikroorganismer har studerats med avseende på parametrar som är, i förhållande till andra parametrar, relativt lätta att identifiera för en fullskaleanläggning; uppehållstiden i akvifären, avståndet mellan infiltrationsbassäng och uttagsbrunn samt hur fort transporten sker mellan bassäng och uttagsbrunn. Resultatet visar att det, trots en begränsning i genomförda studier, går att urskilja vissa trender. Studien visar att avstånd kan vara be­gränsande för ”snabba” akvifärer och en längre uppehållstid för de långsamma. Ett förslag på en metod som skulle kunna användas utifrån analysen av fältstudierna är att använda para­ metrarna transportsträcka, uppehållstid och grundvattenhastighet enligt följande: Tabell 8‑2. Förslag på dimensionering, minimal uppehållstid och avstånd som krävs för reduktion av olika grad. Långsam (1–3 m/dag) Uppehållstid (dygn) Avstånd (m) Snabb (>25 m/dag) Uppehållstid (dygn) Avstånd (m) Erhållen Reduktion (log10 ) >5 10 ca 1 25 3 >14 15 ca 1,5 40 4 >25 20 ca 2 80 5 >35 30 ca 3 >80 6 40 grus oftast i anslutning till isälvsavlagringar, borde en kort uppehållstid (eller egentligen den snabba strömningen) kompenseras med ett längre avstånd. I Sverige är förutsättningarna lite sämre med tanke på att mineralkompositionen ofta är de som reducerar mikroorganismer sämst (Flynn, Rossi & Hunkelers 2004), å andra sidan är koncentrationen av patogener i svenska sjöar och vattendrag generellt lägre (se tabell 3-2). Kalkning sker i flera av de svenska anläggningarna (Jönsson & Wikström 2003) för att öka pH, alkalinitet och hårdhet. Kalk inlagd i infiltrationsbassängerna skulle kunna påverka reduktionen av mikroorganism­ erna och borde undersökas. pH ökar vilket skulle kunna medföra en sämre mikrobiologisk barriär vid infiltration. Flynn, Rossi och Hunkeler (2004) fann dock i sina försök att kalkstenssand var effektiv för fastläggning av mikroorganismer. Det är även andra material som skulle kunna vara aktuella för att agera som ett komplement till sanden i infiltrationsbass­ ängerna, bl.a. järnoxidtäckt olivinsand, vilket behöver studeras noggrannare tillsammans med andra materialskillnader. Andra operativa åtgärder som skulle kunna vara intressanta att studera är pH-just­ ering och salt/jonstyrkejustering och hur de påverkar reduktionen av mikroorganismer. Biohudens betydelse för processen konstgjord grundvattenbildning är inte utredd mer än att den är av stor betydelse för processens effektivitet. I den studerade litteraturen har det inte funnits något försök som undersöker hur biohuden fungerar, vad som skulle kunna påverka dess effektivitet eller en kvalitativ eller kvantitativ bedömning av hur den optimeras, vilket skulle kunna vara ett intressant område för utveckling. En ny grundläggande inventering av ett urval av svenska anläggningar för konstgjord grundvattenbildning borde genomföras som en uppdatering på Sundlöf och Kronqvists studie (1992). Det har skett en del forskning och utveckling på området det senaste decenniet, som skulle kunna komma befintliga och nya anläggningar till del om en ny nulägesbeskrivning genomförs. I denna borde en under­ sökning göras av den mikrobiologiska barriärverkan vid de olika anläggningarna. Det skulle även vara intressant att studera råvattnet för olika vattenverk med speciell fokus på virusförekomsten. Relationen mellan inaktivering och avskiljning är ett område som kräver ytterligare forskning. Virus är så pass okänsliga och har hög överlevnad både i 9 Forsknings- och utvecklingsbehov Med utgångspunkt från den här litteraturstudien kan det konstateras att det finns ett forskningsbehov inom området mikrobiologisk barriär vid konstgjord grund­ vattenbildning. Studien ger en tydlig bild av vilka parametrar som inverkar och hur, men deras samman­ lagda betydelse kan bara studeras i fullskaliga under­ sökningar. Denna typ av försök har inte genomförts i Sverige och som tidigare diskuterats är det, med tanke på olika rådande förhållanden, bäst att studera barriärverkan i så liknande och närliggande miljö som möjligt. För att kunna göra en korrekt bedömning av en anläggning eller dimensionera en framtida anläggning, finns behov av en specifik metod som skulle kunna utgå ifrån resonemanget i kvantifieringsstudien och utvecklas med fler faktorer som påverkar reduktionen, t.ex. omättad zon, koncentration av patogener i råvattnet, material och porositet i akvifären etc. En sådan metod skulle kunna kompletteras med kriterier och en allmän instruktion för att undersöka och av­ göra om ett råvatten är ytvattenpåverkat på en anläggning. Generella regler måste alltid kompletteras med avvägningar grundade på lokala miljöförhåll­ anden, liksom egenskaper hos det vatten man önskar infiltrera (Stenström 1980). En mängd studier har undersökt virusadsorption på fasta ytor under de två–tre senaste decennierna och mycket kunskap har samlats in. Men generella slutsatser om graden av adsorption och mekanismerna som är involverade är svåra att dra för olika typer av ytor under olika kemiska förhållanden. Det beror på att ingen standardiserad metod eller systematiskt angreppssätt har blivit utvecklad och följd (Zhuang & Jin 2003). Även fullskaleförsöken borde kunna homogeniseras så det blir enklare att jämföra result­ aten och genomföra ovanstående process. Kunskap om flera parametrar krävs för att säkerställa en reduktion som uppnår eftersträvt värde och det bästa är att göra noggranna undersökningar på plats. Därför skulle det vara intressant att utföra fler medelstora pilotförsök och studera parametrarna som påverkas av konstruktion och drift av en anläggning. Eftersom materialet som används vid konstgjord grundvattenbildning i Sverige mest liknar materialet i de ”snabba” akvifärerna i studien, dvs. lite större 41 grundvatten och i mark att det inte går att dimens­ ionera en anläggning utifrån inaktiveringstid för enteriska virus. Om dimensionering av anläggningar för konstgjord grundvattenbildning utgår ifrån endast avskiljning sker borde ett säkert intervall uppnås efter­ som även inaktivering bidrar till den totala redukt­ ionen. 42 Referenser Auckenthaler A., Raso G., Huggenberger P., 2002. Particle transport in a karst aquifer: natural and artificial tracer experiments with bacteria, bacteriophages and microspheres. Water Science and Technology 46, 3, 3609–3614. Bengtsson, G., 2005. Seminarieanteckningar från Seminarium om konstgjord grundvattenbildning, Svenskt vatten, Växjö, 2005-10-19. Bouwer H., 2002. Artificial recharge of groundwater: hydrology and engineering, Hydrology Journal (2002) 10:121–142, Springer-Verlag. Chattopadhyay S., Puls R.W., 1999. Adsorption of bacteriophages on clay minerals. Environmental Science and Technology, 33, 3609–3614. Chu Y., Jin Y., Baumann T., Yates M.V., 2003. Effect of Soil Properties on Saturated and Unsaturated Virus Transport through Columns. Journal of Environmental Quality 32, 2017–2025. DeBorde D., Woessner W.W., Kiley Q.T., Ball P. 1999. Rapid transport of viruses in a floodplain aquifer, Water Research 33, 2229–2238. Dizer H., Grützmacher G., Holzbecher E., Szewzyk R., Lopez-Pila J.M., 2005. Elimenation of Coliphages and E.coli by slow sand filtration. Poster på ISMAR 5 Konferens Berlin, 12–16 juni 2005. Djurberg H., 2005. Sprinklerinfiltration – erfarenheter från Katrineholm. Sem­ inarieanteckningar från Seminarium om konstgjord grundvattenbildning, Svenskt vatten, Växjö, 2005-10-19. Ederth T., 2005. Föreläsningsanteckningar TFKI30 Yt- och kolloidkemi, Kap. 5 Fast yta/gas-gränsen, http://www.lith.liu.se/tfk/miklavcic/forelasningar/kap5. pdf (2005-11-02). EPA, United States Environmental Protection Agency, 1993. Preventing Water­ borne Disease, A focus on EPA’s research. EPA/640/K-93/001. Flynn R.M., Rossi P., Hunkeler D., 2004. Investigation of virus attenuation mechanisms in a fluvioglacial sand using column experiments. FEMS Microbiology Ecology 49, 83–95. Foppen J.W.A., Mporokoso A., Schjiven J.F., 2005. Determining straining of Escherichia coli from breakthrough curves. Journal of Contaminant Hydrology 76, 191–210. 43 Fryklund C., 1992. Artificial Groundwater Recharge – state of the art. VA-Forsk Rapport 1992-04. Fryklund C., Jacks G., Johansson P-O., Lekander K., 1994. Konstgjord grund­ vattenbildning – processtudier vid inducerad infiltration och bassänginfiltration. VA-Forsk Rapport 1994-08. Guan H., Schulze-Makuch D., Schaffer S., Pillai S., 2003. The effect of critical pH on virus fate and transport in saturated porous medium. Ground Water 41, 5, 701–708. Göteborgs va-verk, 2004. Vad betyder parametrarna? www.vaverket.goteborg.se >Dricksvatten>Råvattenkvalitet (2005-08-12). Göteborgs va-verk, 2005. Vattenkvalitet i Göta älv 2004 (Jan–Dec) www.vaverket. goteborg.se >Dricksvatten>Råvattenkvalitet> Sammanställning av analysresultat: Göta älv, Lärjeholm kvartal 1–4/2004 (2005-10-10). Hanson G., 2000. Konstgjord grundvattenbildning: 100-årig teknik inom svensk dricksvattenförsörjning, VA-Forsk Rapport 2000-5. Havelaar A.H., van Olphen M., Schjiven J.F., 1995. Removal and inactivation of viruses by drinking water treatment processes under full scale conditions. Water Science and Technology 31, 55–62. Hijnen W.A.M., Schijven J.F., Bonné P., Visser A., Medema G.J., 2004. Elimination of viruses, bacteria and protozoan oocysts by slow sand filtration. Water Science and Technology 50 No1, 147–154. HRV Water Purification AB, 2005. Mikroorganismer i vattnet http://www.hvr.se/ vatten/mikroorganismer_i_vatten.html (2005-07-04). Jensen T.Ø., Berg R.L., Bennedsen L., Brandt G., 2005. Geochemical and micro­ bial processes in the unsaturated zone at the Arrenaes artificial recharge trial plant. Bidrag till ISMAR 5 konferensen Berlin, 12–16 juni 2005. Jin Y., Yates M., Thompson S., Jury W., 1997. Sorption of viruses during flow through saturated sand columns. Environmental science and technology 31, 548–555. Jin Y., Chu Y., Li Y., 2000. Virus removal and transport in saturated and unsaturated sand columns. Journal of Contaminant Hydrology 43, 111–128. Jönsson R., Wikström A-S., 2003. Vattenbehandling genom återinfiltration i filter­ bäddar med skikt av krossad kalksten. Va-Forsk Rapport 2003-06. Jörgensen C., Peters J., Eschweiler B., 2001. Removal of pathogens and monitoring of microbial changes during artificial recharge. Artificial recharge of groundwater, EC project ENV4-CT95-0071 Final report, Kapitel 8. 44 Karlstad kommun, 2005. Vattnets väg. http://www.karlstad.se/tv/va/vatten/ vattnetsvag2.shtml (2005-02-23). Leclerc H., Edberg S., Pierzo, V., Delattre J.M., 2000. Bacteriophages as indicators of enteric viruses and public health risks in groundwater. Journal of Applied microbiology 88, 5–21. Livsmedelsverket, Vägledning till Livsmedelsverkets föreskrifter om dricksvatten, 2004-05-01, www.slv.se (2005-10-24). Macler B., Merkle J.C., 2000. Current knowledge on groundwater microbial pathogens and their control. Hydrogeology Journal 8, 29–40. Niemi R.M., Kytövaara A., Pääkkönen J., Lahti K., 2004. Removal of F-specific RNA bacteriophages in artificial recharge of groundwater. Water Science and Technology 50, 1, 155–158. Ottoson J., Stenström T.A., 2003. Faecal contamination of greywater and associated microbial risks. Water Research 37, 645–655. Pang L., Close M., Goltz M., Sinton L., Davies H., Hall C., Stanton G., 2003. Estimation of setback distances based on transport of E.coli and F-RNA phages. Environment International 29, 907–921. Pang L., Close M., Goltz M., Noonan M., Sinton L., 2005. Filtration and trans­ port of Bacillus subtitles spores and the F-RNA phage MS2 in a coarse alluvial gravel aquifer: Implications in the estimation of setback distances. Journal of Contaminant Hydrology 77, 165–194. Persson, F., Heinicke, G., Hedberg, T., Bergstedt, O., Wångsell, C., Rydberg, H., Kjellberg, I., Kristenson, S-E., 2005. Mikrobiologiska barriärer i vattenrening. Va-Forsk Rapport 2005-17. Peters J.H, van Baar M.J.C., Nobel P.J., Vogelaar A.J., Schijven J.F., Hoogen­ boezem W., Bergsma J., Stakelbeek A., 1998. Fate of pathogens and consequences for the design of artificial recharge systems. I Peters et al. (red.) Artificial recharge of groundwater. Proceedings of the third international symposium on artificial recharge of groundwater – TISAR 98, Amsterdam, s. 141–148. Prescott L.M., Harley J.P., Klein D.A., 1999. Microbiology, fourth edition. WCB/ McGraw-Hill. Rydberg H., 2005. Labchef, Lackarebäcks laboratorium, Göteborgs vatten och avloppsverk, Personlig kontakt, 2005-09-13. Rzeżutka A., Cook N., 2004. Survival of human enteric viruses in the environment and food. FEMS Microbiology reviews 28, 441–453. 45 Schijven J.F., Medema G., Vogelaar A.J., Hassanizadeh S.M., 2000. Removal of microorganisms by deep well injection. Journal of Contaminant Hydrology 44, 301–327. Schijven J.F., 2001. Virus removal from groundwater by soil passage, Modelling, field and laboratory experiments. Ponsen & Looijen B.V, Wageningen. Schijven J.F., Hassanizadeh S.M., de Bruin H.A.M., 2002. Two-site kinetic modelling of bacteriophages transport through columns of saturated dune sand. Journal of Contaminant Hydrology 57, 259–279. Schijven J.F., de Bruin H.A.M., Hassanizadeh S.M., de Roda Husman A.M. 2003. Bacteriophages and clostridium spores as indicator organisms for removal of pathogens by passage through saturated dune sand. Water Research 37, 2186–2194. Schäfer A., Ustohal P., Harms H., Stauffer F., Dracos T., Zehnder A.J.B., 2004. Transport of bacteria in unsaturated porous media. Journal of Contaminant Hydrology 33, 149–169. Smittskyddsinstitutet, 2005. Vattenburen smitta. http://www.smittskyddsinstitutet.se/SMItemplates/Article____2376.aspx (2005-08-12). Stenström T.A., 1980. Reduktion av bakterier och virus vid avloppsinfiltration i mark – en kunskapssammanställning, SNV pm 1329. Stenström T.A., 1985. Infiltration i mark – Mikroorganismers transport och överlevnad, SNV pm 3051. Stevik T-K., Aa K., Ausland G., Hanssen J.F., 2004. Retention and removal of pathogenic bacteria in wastewater percolating through porous media: a review. Water Research 38, 1355–1367. Strand M., 2005. Processingenjör, Karlskrona Vattenverk, Råvattenkvalitet Karlskrona, personlig kontakt 2005-10-24. Stockholm Vatten 2005. Vattenkvalitet vid Lovö vattenverk. www.stockholmvatten.se > Dricksvatten > Vattenkvalitet (2005-11-01). Sundlöf B., Kronqvist L., 1992. Konstgjord grundvattenbildning, Nulägesbeskriv­ ning av tjugo svenska anläggningar. Svenska vatten och avloppsföreningen, VA-Forsk Rapport 1992-13. Thompson S., Flury M., Yates M., Jury W., 1998. Role of the Air-Water-Solid Interface in Bacteriophage Sorption Experiments. Applied and Environmental Microbiology, Jan, 304–309. Thougaard H., Varlund V., Madsen R.M., 2001. Grundläggande mikrobiologi med livsmedelsapplikationer. Studentlitteratur, ISBN: 9144015690. 46 Wall K., Toze S., O’Hara G., 2005. Interactions of ondigenous groundwater bacteria with enteric viruses during water quality improvement by aquifer storage and recovery (ASR). Poster på ISMAR 5 Konferens Berlin, 12–16 juni 2005. WHO, World Health Organization, 2004. Guidelines for drinking-water quality, third edition. http://www.who.int/water_sanitation_health/dwq/gdwq3/en/index.html, 200507-04. Wikipedia, 2005. www.wikipedia.com. Sökord Protozoa och Bacteria (200508-12). Zhuang J. och Jin Y., 2003. Virus retention and transport through Al-oxide coated sand columns: effects of ionic strength and composition. Journal of Contaminant Hydrology 60, 193–209. 47 Bilaga A: Sökord Artificial recharge Infiltration Groundwater Aquifer Microbial contamination Microbial transport Indigenous groundwater bacteria Virus transport Filtration (Ad)Sorption Inactiviation Column experiments Sand column Saturated transport Unsaturated transport Saturated zone Unsaturated zone Reduction Survival Microorganisms Pathogen Protozoa Bacteria Virus Bacteriophages MS2 phiX174 PRD1 E.coli Minerals Gravel, sand 48 49 Havelaar 1995 Erskine, USA Undersökn.omr. Deborde 1999 FRNA Enterovirus Tot koliformer Inducerad infiltration Sand 0,2–0,24 mm Polio X174 PRD1 MS2 Mikro­ organism Remmerden, Rijn (Rhen) Öppen akvifär, grovt grus och sten (porositet 0,15) som övergår till sand under 6 m. Plats, typ av akvifär och material Referens 30 30 30 17,24 19,4 35,56 40 6,67 26,67 30 7,5 17,24 19,4 35,56 40 6,67 26,67 30 7,5 17,24 19,4 35,56 40 6,67 26,67 30 7,5 17,24 6,67 Uppe­hållstid (tim) 19,4 7,5 Avstånd (m) Bilaga B: Data från fältstudier 14 0,72 0,28 1,48 1,11 0,72 0,28 1,48 1,11 0,72 0,28 1,48 1,11 0,72 0,28 Uppe­hållstid (dygn) 5 4 5 5,0 4,0 7,5 6 4,0 2,5 6 5 3,5 2,5 6 5 3,5 2,5 Avskiljning -log(c/c0) 0,17 0,13 0,17 0,26 0,53 0,19 0,20 0,21 0,33 0,15 0,17 0,18 0,33 0,15 0,17 0,18 0,33 Reduktionsfaktor (-log(Cmax/c0)/m) ca 2,2 ca 2,2 ca 2,2 27 27 27 27 27 27 27 27 27 27 27 27 27 27 v (m/ d) 1,13 1,13 1,13 1,13 1,13 1,13 1,13 1,13 1,13 1,13 1,13 1,13 1,13 1,13 v (m/h) 105 –106/l 102/l Ca 103 –104 /l 3,4·106 2,9·107 5,4·109 5,6·1010 c0 (pfu/ml) 50 Pang 1998 ref i Pang 2005 Sinton 2000 ref i Pang 2005 Castricum, Nederländerna Pang 2003 & Schjiven 2001, s 75 Burnham 1998 Heterogent material, grus 0,15 mm samt stråk av lera Burnham, Nya Zeeland Fin sand ca 98 %, 1,5 %silt och 0,5 % lera, medel partikelstorlek 0,2– 0,24 mm (porositet 0,35 Plats, typ av akvifär och material Referens B. Subtitlis MS2 FRNA (PRD1 och MS2) Mikro­ organism 12,0 26 17 30 9,71 11,32 10,68 19,68 21,38 30,07 25,36 36,8 37,6 61,6 61,8 86 87 18,67 87 20 16,89 61,8 6,33 9,62 37,6 19,8 8,27 8,60 36,4 36,8 1,06 1,25 0,89 0,82 0,45 0,47 0,40 0,26 0,78 0,70 0,40 0,36 0,34 0,20 7,0 10 20,6 4,0 6,4 1,5 Uppe­hållstid (dygn) 2,5 4,70 Uppe­hållstid (tim) 3,8 2,4 Avstånd (m) 6,2 8,4 5,5 5,7 4,6 4,7 8,1 5,5 5 4,7 3,9 3,9 4,2 3,3 8,4 6,5 5,6 4,9 3,3 3 Avskiljning -log(c/c0) 0,07 0,10 0,09 0,09 0,12 0,13 0,41 0,28 0,06 0,08 0,10 0,11 0,12 0,16 0,28 0,38 0,56 0,77 0,87 1,25 Reduktionsfaktor (-log(Cmax/c0)/m) v (m/h) 82,32 68,64 69,36 75,12 84,48 78 49,44 75,12 111,84 87,84 93,84 102,72 105,6 105,12 3,43 2,86 2,89 3,13 3,52 3,25 2,06 3,13 4,66 3,66 3,91 4,28 4,40 4,38 räknat på medel: 1,39 1,19–1,59 v (m/d) 2,2·1012 resp. 2,2·1013 c0 (pfu/ml) 51 Kokemäenjoki, Finland Niemi 2004 Schijven 2000 Burnham Sinton 2000 ref i Pang 2005 E.coli R5 (Cl.per) PRD1 25 38 22 38 2,4 6,1 12 8 2,4 25 38 22 38 8 6,1 12 38 2,4 912 25 6,1 2,4 19 12 5 0,08 0,69 0,44 0,31 Uppe­hållstid (dygn) 8 38 600 146,4 12 22 456 24 Djup-infiltration 288 17 Sand, fluvial sediment, Fin sand ca 98 %, 1,1 % silt och 0,9 % lera, medel partikelstorlek 0,27 mm (porositet 0,32) 120 11 2 10,59 16,52 37,6 61,8 5 7,35 Uppe­hållstid (tim) 36,8 Avstånd (m) 57,6 MS2 MS2 B. Subtitlis Mikro­ organism 8 Someren, Nederländerna Grov sand Plats, typ av akvifär och material Referens 7,5 5,2 5 4,4 4,7 7,1 5,7 8,4 8 7,2 5,9 7 6,9 3,3 1,5 5,4 4,5 4,1 Avskiljning -log(c/c0) 0,94 0,14 0,23 0,37 0,59 0,59 0,71 0,22 0,36 0,60 0,74 0,29 0,41 0,30 0,30 0,09 0,12 0,11 Reduktionsfaktor (-log(Cmax/c0)/m) 3,3 1,01 0,9 1,98 3,3 1,01 0,9 1,98 3,3 1,01 0,9 1,98 3,3 1,26 1,42 2,2 60 89,76 85,2 120,24 v (m/d) 0,05 0,06 0,09 2,5 3,74 3,55 5,01 v (m/h) 1012/l 2·1010 /l 5·1011/l 7·1013 /l c0 (pfu/ml) Mikrobiologisk barriärverkan vid konstgjord grundvattenbildning – en litteraturstudie om påverkande faktorer Box 47607 117 94 Stockholm Tfn 08 506 002 00 Fax 08 506 002 10 E-post [email protected] www.svensktvatten.se