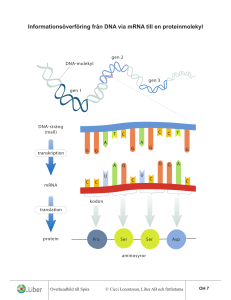
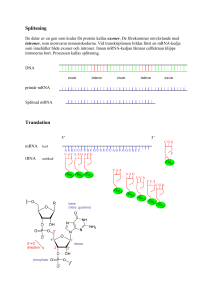
SVAR: 1.a) Förslag: Gelfiltreringskromatografi, kommer att ge 4
advertisement
 Förslag: Gelfiltreringskromatografi, kommer att ge 4")
SVAR: 1.a) Förslag: Gelfiltreringskromatografi, kommer att ge 4 baslinjeseparerade toppar. Elueringsordning störst först, så topp 1 Protein E, topp 2 protein D, topp 3 protein C och B, topp 4 protein A. För att separera protein B och C, jonbyteskromatografi. Katjonbytare, proteinerna binds in vid pH 6,5. Vid detta pH är protein B plusladdat och fastnar i kolonnen, protein C minusladdat och rinner rakt igenom. För att eluera B höj jonstyrkan på bufferten. Kromatogrammet bör endast innehålla en topp. b) Har en reaktiv serin i aktiva ytan. c) Kymotrypsin klyver C-terminalt om stora opolära sidokedjor. Peptiderna innehållande Phe och Trp kan fungera som substrat. d) Kortfattat svarsförslag för fullständigt se fig i Stryer. De 3 aa i aktiva ytan är: Asp, His och Ser. His fungerar som protonrelä (omväxlande bas/syra). Initialt tar His en proton från Ser som då blir en bra nukleofil. Asp håller His på plats och underlättar His roll som protonacceptor. Efter klyvning av peptidbindninen donerar His en proton till aminogruppen. Under enzymregenereringsstegen av katalysen fungerar His i tur och ordning som proton-acceptor och sedan donator. 2a) Approximationen innebär att [ES] är i det närmaste konstant under en viss tid då det fortfarande finns ett överskott på S över E då gäller: d ES 0 k1 E S k 2 ES k 1 ES dt b) kcat beskriver enzymets effektivitet och anger hur mycket substrat som omvandlas till produkt per enzymenhet och per tidsenhet c) 1 ((l*min)/mmol) v 0.45 0.4 Linjens ekvation y = 0.4062x + 0.1976 y = 0.4062x + 0.1976 0.35 0.3 Skärning med y-axel = 0.1976 = 1/vmax Linjens lutning = 0.4062 = KM/vmax 0.25 Series1 0.2 Linear (Series1) 0.15 0.1 0.05 0 0 0.1 0.2 0.3 0.4 0.5 0.6 1 S (mM-1) Beräkna vmax: vmax = 1/0.1976 = 5.06 mmol/(l*min) Beräkna KM: KM = vmax * 0.4062 = 5.06 * 0.4062 = 2.05 mM e) Röd linje visar hur grafen förändras i närvaro av en nonkompetitiv inhibitor. 1 ((l*min)/mmol) v 0.45 KM påverkas inte vmax minskar med faktorn 1+[I]/KI (där KI = [I][E]/[EI]) 0.4 y = 0.4062x + 0.1976 0.35 0.3 0.25 Series1 0.2 Linear (Series1) 0.15 0.1 0.05 0 0 Linjen lutning (= KM/vmax) ökar i närvaro av inhibitorn Skärningen med y-axeln (= 1/vmax) ökar i närvaro av inhibitorn Skärningen med x-axeln (=-1/KM) förändras inte 0.1 0.2 0.3 0.4 0.5 0.6 1 S (mM-1) 3. a) Fosfatgrupperna pekar utåt och kvävebaserna pekar inåt. b) En fosfatgrupp kopplar två sockerenheter via ett syre på 3’-kolet och ett syre på 5’kolet där 3’betyder kolatom nummer 3 i sockerenheten och 5’ betyder kolatom nummer 5 i sockerenheten. c) Ny trifosfatnukleotid binder först till mallkedjan genom basparning i.e. vätebindning mellan två matchande kvävebaser Hydroxylgruppen på C3 i den intilliggande nukleotiden gör en nukleofil attack på den innersta fosfor-atomen i den nya trifosfatnukleotiden. Pyrofosfat (PPi) hydrolyseras och den nya nukleotiden sätts ihop kovalent med resten av DNA kedjan Energikrävande process - energi frigörs då PPi hydrolyseras d) Om ett fel uppstår vid replikationen och inte åtgärdas kommer felet att finnas kvar hos kommande generationer av celler vilket kan resultera i permanenta fel vid syntes av proteiner. Ett fel vid transkriptionen leder till ett felaktigt mRNA (och fel vid proteinsyntesen) men eftersom mRNA till slut bryts ned (då proteinet inte längre skall syntetiseras) så blir felet inte permanent. DNA-polymeras I har s.k. exonukleasfunktion och det innebär att den kan hydrolysera bort en felaktigt inkorporerad nukleotid. e) Om RNA innehåller två efterföljande sekvenser med många G och C så kan dessa baspara med varandra och bilda en stabil hårnålsloop (G och C basparar med tre vätebindningar). Om det sedan kommer en sekvens på RNA som innehåller många U så sker en destabilisering i interaktionen mellan RNA och DNA (U och A basparar med två vätebindningar) vilket leder till att RNA:t lossar från DNA och RNA-polymeraset släpper. Ett annat sätt att avsluta transkriptionen sker genom proteinet rho som känner igen en sekvens i det nysyntetiserade transkriptet och binder till RNA:t vilket leder till att RNA-polymeraset lossnar. Processen kräver energi (hydrolys av ATP → ADP + Pi). 4. a) De primära mRNA transkripten i eukaryoter består av exoner och introner. Exonerna innehåller den genetiska informationen som kodar för protein. Intronerna används inte som mall för proteinsyntes utan dessa klipps bort från mRNA – processen kallas för splicing. Ofta kodar exonerna för olika funktionella domäner eller subenheter i ett protein. Genom alternativ splicing kan olika varianter av mRNA bildas från en och samma kodande region på DNA – detta gör att vi kan producera betydligt fler varianter av mRNA än det antal gener vi har. Ex. en gen som innehåller fyra exoner (nummer 1, 2, 3 och 4). Mellan exonerna finns introner. När intronerna skall klippas bort från mRNA kan detta ske på olika (alternativa sätt) vilket innebär att en eller flera exoner också tas bort. På så vis kan olika varianter av mRNA bildas (ex. mRNA med exon 1, 2 och 4 eller mRNA med exon 1,3 och 4 osv). b) Ett kodon är nukleotidtriplett i mRNA som kodar för en viss aminosyra vid proteinsyntesen. Antikodonet är en nukleotidtriplett i tRNA som vätebinder komplementärt och antiparallellt till mRNA vid proteinsyntesen. Gör att rätt aminosyra byggs in i proteinet. c) Vid start av translationen sitter en tRNA-molekyl med den växande polypeptidkedjan (eller fmet vid första translationsrundan) i P-sitet. En ny tRNA-molekyl med kopplad aminosyra binder in i A-sitet där det sker en matchning mellan kodonet på mRNA i A-sitet och antikodonet på den inkomna tRNA-molekylen. Den växande polypeptidkedjan (eller fmet) på tRNA:t som sitter i P-sitet överförs till aminosyran i A-sitet och en ny peptidbindning bildas. Därefter sker en translokering vilket innebär att den fria tRNA-molekylen i P-sitet flyttar till Esitet, tRNA som binder den växande polypeptidkedjan flyttar från A-sitet till P-sitet och mRNA matas fram motsvarande en bastriplett så ett nytt kodon exponeras i A-sitet. Den fria tRNA-molekylen lämnar E-sitet och en ny translationsrunda kan ske.