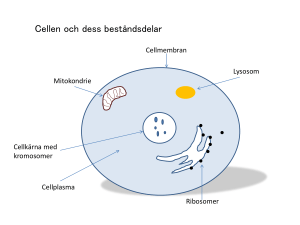
Cell- och molekylärbiologi
advertisement

Välkommen till cellbiologin! (Åbo akademi) Cytoskelettet i en levercell (D. Toivola) Cellen utgör den grundläggande byggstenen för allt liv på jorden. Att känna till hur celler fungerar är därför av väsentlig betydelse för all biologi. Inom cellbiologin studeras hur celler lever och varför de dör, deras livscykel och beteende, deras differentiering till vävnader och organ samt olika störningar i dessa processer. Signalering mellan och inom celler utgör också ett viktigt tema inom cellbiologi liksom förhållandet mellan omgivningen och avläsningen av arvsmassan. I skolningen poängteras även hur störningar i cellers funktioner till olika slags sjukdomar samt hur cellulära processer och styrmekanismer kan utnyttjas för farmakologisk produktutveckling och diagnostik. Modern biologi Biologin befinner sig i ett dynamiskt utvecklingsskede. Från att ha varit begränsad till zoologi, botanik och genetik har den utvecklats till en central del av biovetenskaperna, och anknyter till bl.a. biomedicin, bioteknologi och miljöproblematik. Inom cellbiologin undersöks cellernas och vävnadernas struktur, differentiering och utveckling, samt störningar i dessa. Signalering mellan och inom celler är ett viktigt forskningsområde som ligger mellan cellbiologi och fysiologi. Forskning inom fysiologi undersöker cellers och vävnaders funktion och störningar i dessa. Biologin vid Åbo Akademi har strävat till att utveckla tekniker som ger möjlighet att bedriva högklassig cellbiologisk forskning. Stora anskaffningar har varit konfokal- och tvåfotonmikroskop, apparatur för dynamisk fluorimetri, apparatur för mikroinjicering i levande celler samt etablerandet av en utvecklingsbiologisk enhet för produktion av sebrafiskembryo. ÅA Biologin har även ett transmissionselektronmikroskop. Förutom dessa metoder bedrivs en omfattande molekylbiologisk och proteinkemisk analys av signalmolekyler i celler. Vi har även tillgång till en omfattande apparaturpark i Bioteknikcentret. Ämnespresentation Immunceller som diskuterar (T. Söderström) Cellbiologin utgör en central del av biovetenskaper, biomedicin, bioteknologi och anknyter även till ekologi och miljöproblematik. Tack vare den fenomenala utveckling som skett inom cellbiologin under de senaste årtiondena utgör cellbiologins tekniker och koncept helt centrala redskap inom alla biovetenskaper och sammanhänger även med de raska framsteg som skett inom t.ex. stamcellforskning, läkemedelsframställning, diagnostik samt bio-nanoteknologisk forskning. Cellbiologiundervisningen ger en solid kunskapsbakgrund i cellbiologins alla delområden samt även i molekylär genetik, cellsignalering, molekylärbiologi, immunologi, fysiologi, djuranatomi och även goda insikter i biokemi och kemi. Syftet är att ställa till förfogande hela paletten av de modernaste tekniker som krävs att bedriva högklassig cellbiologisk forskning. Olika slags avancerade mikroskopiska och andra biovisualiseringsteknologier är en viktig del av undervisningen, vilket sker i samarbete med Medicinska fakulteten vid Åbo universitet samt Åbo universitetsjukhus. För att trygga tillgången till toppmodern utrustning av världsklass har omfattande anskaffningar på cellvisualiseringens område nyligen verkställts och denna utveckling kommer att fortsätta. Cellbiologin kan även utnyttja det kursprogram som finns till förfogande inom det nya internationella magistersprogram med rubriken"Masters program in biomedical imaging" som inleds hösten 2010. Förutom visualiseringsmetoder bedrivs en omfattande molekylärbiologisk och proteinkemisk analys av signaloch transkriptionsmolekyler i celler. Cellbiologin har även tillgång till en omfångsrik apparaturpark i Bioteknikcentret. ( www.btk.fi). Molekylärbiologi (Wikipedia) Molekylärbiologi är en disciplin inom biologin som söker svar på frågorna om hur biologin fungerar på molekylär nivå. Fältet överlappar andra områden inom biologi och kemi, särskilt genetik och biokemi. Molekylärbiologin sysslar huvudsakligen med hur olika molekyler och system inuti cellen interagerar, och särskilt med flödet av genetisk information från DNA via RNA till proteinsyntes, samt hur dessa processer regleras. Molekylärbiologin intresserar sig mycket för de biologiska molekylernas form, men sysslar också med hur de blir till och med deras funktion. Molekylärbiologin blev en etablerad vetenskapsgren under 1930-talet. Termen molekylärbiologi myntades dock först 1938 av Warren Weaver, som var chef för stiftelsen Rockefeller Foundation i New York. Han antog att biologin stod inför en period av stora förändringar, till följd av tekniska framsteg såsom röntgenkristallografi och de möjligheter som detta medförde. Därför såg han till att avsevärda penningsummor tillfördes det biologiska fältet. Molekylärbiologins relation till besläktade vetenskaper Schematisk skiss över förhållandet mellan biokemi, genetik och molekylärbiologi. Forskare inom molekylärbiologi använder metoder och tekniker som ursprungligen hör hemma inom molekylärbiologin, men dessa kombineras i allt större utsträckning med tekniker och idéer från genetik, biokemi, och biofysik. De tydliga gränser som en gång fanns mellan dessa områden har suddats ut. Figuren ovan vill visa ett sätt att se de olika disciplinernas relation till varandra. Biokemin studerar de kemiska substanser och viktiga processer som sker i levande organismer. Genetiken studerar vilken effekt genetiska skillnader har på organismen. Ofta kan man dra slutsatser om detta genom att jämföra individer som saknar en viss gen med individer vars motsvarande gen är fullt funktionell. Ofta studerar man muterade individer som saknar en eller flera viktiga gener jämfört med den så kallade vildtypen, den normala fenotypen. Detta kan göras i bakterier såväl som i möss. De enkla slutsatserna av dessa studier störs dock ofta av att olika gener kan reglera uttrycket av en annan gen, och av att många gener har olika funktioner under fosterstadiet och hos den vuxna individen. Molekylärbiologin studerar den molekylära basen för överföring av genetisk information. Molekylärbiologin studerar främst replikationen, då kromosomer kopieras vid celldelning, transkriptionen då informationen i DNA översätts till RNA, och translationen då den genetiska informationen når sitt slutresultat i form av tillverkning av protein. Centrala dogmen säger att flödet av genetisk information går från DNA via RNA till protein. Denna bild har visat sig förenklad, inte minst genom upptäckten av retrovirus, men även genom upptäckten av flera funktioner hos RNA. Även om dogmen inte är heltäckande förblir den ändå basen och inkörsporten till fältet. En stor del av det molekylärbiologiska arbetet är kvantitativt - man vill inte bara veta om något finns eller inte, utan även hur mycket. Under 2000-talets första år har mycket hänt vad gäller användandet av datorer inom bioinformatiken, att dra slutsatser av genetiska data samlade i gigantiska databaser. Molekylär genetik har klivit fram som det viktigaste fältet inom molekylärbiologin. Fler och flera fält inom biologins studerar även den molekylära nivån. Inom cellbiologin och utvecklingsbiologin studerar man molekylära interaktioner i sig. Inom populationsgenetik, där man studerar genetiken i större populationer och inom fylogenetiken, som studerar hur olika arter är släkt med varandra, studeras biomolekylerna indirekt då de molekylärbiologiska metoderna används för att dra slutsatser. Även inom biofysiken finns en lång tradition av att studera biomolekylernas basala egenskaper. Väsentliga molekylärbiologiska tekniker En PCR-maskin. Enkelt uttryckt består denna av ett programmerbart värmeblock, som ändrar temperatur i en cykel om tre steg och sedan upprepar denna så många gånger man önskar. Sedan 1950-talet och tidigt 1960-tal har molekylärbiologerna lärt sig att karaktärisera (identifiera och beskriva), isolera (renframställa) och manipulera cellernas och organismernas molekylära beståndsdelar. Hit hör DNA, den form som den genetiska informationen lagras i, RNA, en besläktad molekyl vars funktioner sträcker sig från att utgöra en arbetskopia av DNA till att fungera som enzym och att utgöra en viktig del av translationsapparaten, och proteiner som är den viktigaste typen av strukturella molekyler i en cell och dessutom omfattar en stor mängd viktiga enzymer. PCR PCR är en förkortning från engelskans polymerase chain reaction - kedjereaktion med hjälp av ett enzym, polymeras. PCR är troligen den enskilda teknik som mest snabbat på utvecklingen inom molekylärbiologin. Det är en snabb och oerhört mångsidig teknik som man med hjälp av kan kopiera upp DNA-sekvenser. I korthet kan man med PCR tillverka miljontals kopior av några få exemplar av sin gensekvens, genom en upprepad serie av temperaturförändringar där ett enzym för varje cykel upp till fördubblar mängden kopior av sekvensen. Man kan framställa en sekvens som är identisk med ursprungssekvensen, eller förändra den på önskat sätt. Man kan stoppa in sekvenser där restriktionsenzym kan klyva DNA-sekvensen, eller mutera sekvensen. PCR kan även användas för att söka efter en sekvens i ett cDNA-bibliotek. Att uttrycka gener Att klona (mångfaldiga) en gen med hjälp av en plasmid. 1. Kromosomalt DNA från den organism man vill studera, organism A 2. Mångfaldigande av en gen med PCR 3. Stor mängd av genen från organism A 4. Genen infogas i en plasmid. 5. Plasmiden med genen från organism A 6. Plasmiden förs in i organism B 7. Överst: genen tillverkas i många kopior av värdcellen. Nederst: värdcellen uttrycker protein med genen som mall. En av molekylärbiologins mest basala tekniker är att uttrycka gener, på engelska expression cloning. För att uttrycka genen stoppas DNA som kodar för det protein man är intresserad av in i en expressionsvektor, till exempel en plasmid. Man säger att man klonar in genen i vektorn. Expressionsvektorn är försedd med promotorelement som driver på produktionen av proteiner ifråga. Oftast har den också gener som kodar för antibiotikaresistens, för att man lätt ska kunna detektera var vektorn finns. Vektorn kan föras in i bakterier eller djurceller. Plasmider kan föras in i bakterier via exempelvis elektroporering, där man använder en kortvarig elchock för att få cellernas membran att öppna sig och släppa in plasmiden. Detta kallas transformation. Att föra in DNA i eukaryota celler, såsom djurceller, kallas transfektion och kan ske med hjälp av exempelvis kalciumfosfatmetoden eller liposomtransfektion. Om DNA istället förs in i cellen med hjälp av ett virus eller en bakterie kallas det för transduktion. När väl det DNA som kodar för den gen man vill studera finns inuti cellen kan den utgöra en mall för proteintillverkning - genen uttrycks. Det finns ett flertal olika system för att se till att cellen producerar höga mängder protein, som sedan kan utvinnas ur cellen. Detta protein kan sedan användas för många olika syften. Man kan studera dess funktion till exempel genom att undersöka dess enzymatiska aktivitet eller studera dess tredimensionella struktur. Läkemedelsindustrin kan använda tekniken för att testa olika potentiella läkemedelsmolekylers effekter på proteinet. Immunohistokemi Immunohistokemi är att använda antikroppar som binder den molekyl man är intresserad av, för att detektera den. Alla immunokemiska metoder är i sig behäftade med en viss osäkerhet, så det är svårt att vara säker på att antikroppen inte binder även till något annat än det man vill studera. Som komplement till andra metoder kan dock immunokemi ge viktig information. I regel används två antikroppar: en som binder till målmolekylen, och en annan som binder till den första antikroppen men också genererar en signal som kan detekteras. Immunokemi för att detektera ett protein på snitt av vävnad från människa eller djur kallas immunohistokemi. Immunohistokemi talar om ifall proteinet finns, och visar var någonstans i vävnaden. Genom att kombinera flera antikroppar som binder olika proteiner kan western blot även tala om vilken celltyp som uttrycker det protein man studerar, och vilka proteiner vars uttryck sammanfaller med forskarens primära målprotein. Antikroppar för immunokemi tillverkas oftast genom att en liten mängd av det protein man vill studera injiceras i ett djur. Därigenom provoceras djurets immunförsvar att tillverka antikroppar mot det främmande proteinet, antigenet. Ofta används kanin eller mus för att framställa antikroppar, men även får och åsna är vanliga. Alternativa metoder för att framställa antikroppar utan att använda djur finns, men tillämpas ännu (2005) endast i mindre skala. Antikroppar kan även användas för att rena proteiner, ofta proteiner som framställts artificiellt i bakterier eller djurceller. Gelelektrofores PCR-produkt körd på agarosgel. Längst till vänster (1) en storleksmarkör, en så kallad stege. Genom att ladda DNA av känd storlek på gelen, kan man bestämma storleken på DNA-fragmenten i provet. bp står för baspar. I mitten (2) provet, PCR-produkt. Till höger (3) samma DNA-fragment, i större mängd. Gelektrofores tillhör molekylärbiologins basala redskap. Tekniken används för att separera molekyler, primärt efter storlek. Man använder sig av ett elektriskt fält, och låter sitt prov vandra genom ett tunt skikt av en porös gel. Små molekyler kommer då att vandra snabbare än stora, och på så sätt separeras molekyler av olika storlek från varandra. För DNA används vanligen agarosgeler, medan proteiner separeras på en gel av polyakrylamid med en teknik som kallas SDSPAGE. Proteiner kan även separeras med avseende på deras elektriska laddning, genom isoelektrisk fokusering. Blottningstekniker De flesta blottningstekniker bygger på att man kan föra över DNA, RNA eller proteiner som separerats exempelvis efter storlek, från den gel som körts i gelelektrofores till ett tunt membran där målmolekylerna fastnar. När molekylerna väl fastnat på membranet kan de undersökas eller användas för olika ändamål. Blottningstekniker brukar generellt sett benämnas med det engelska namnet även på svenska. Southern blot Southern blot används för att fastställa molekylvikten och den relativa mängden av en viss DNA-sekvens. Tekniken har sitt namn efter Edward Southern som utvecklade den. En southern blot består av flera steg. Ett DNA-prov körs på gelelektrofores, ofta efter att först ha fragmenterats (delats upp i mindre bitar) genom klyvning av restriktionsenzym. Sedan överförs DNA-bitarna till ett membran. Där får de binda, hybridisera, till en DNA-sekvens som man märkt in så att den går att detektera, en prob. Sedan man tvättat bort prob som inte bundit till DNA-sekvenserna på membranet detekterar man proben. Den bild som erhålls visar dels den eller de platser där proben bundit, och därigenom hur stor den bit DNA som proben bundit till är, och dels ger signalens styrka ett mått på den relativa förekomsten av DNA-sekvensen. Northern blot Northern blot utförs enligt samma principer som southern blot, men med RNA istället för DNA. Northern blot används för att undersöka mängden av en viss mRNA-molekyl, vilket motsvarar hur mycket genen uttrycks på RNA-nivå. Northern blot är en viktig teknik för att bestämma i hur hög grad en viss gen finns uttryckt i en viss vävnad. Under 2000-talet har dock northern blot fått konkurrens av kvantitativ direktanalyserad PCR, en PCR-baserad teknik med samma syfte. Western blot Western blot på proteiner från celler som genomgår differentiering. Prov tagna vid tre olika tidpunkter (0, 1, 2). Allteftersom cellerna differentierar minskar mängden E-cadherin, medan mängden N-cadherin ökar. Western blot är blottningstekniken utförd på proteiner. Först körs en gelelektrofores, en SDS-PAGE, och proteinerna blottas över till ett membran. Sedan används antikroppar för att detektera proteinerna, och visa var på membranet det sitter vilket visar proteinets storlek. Cellens evolution När det gäller uppkomsten av liv finns många teorier. Skapelseberättelsen är en teori. Vid sidan av den tror man att liv långsamt har utvecklats ur död materia. En teori går ut på att livsformer kommit från rymden t.ex. med meteoriter eller t.o.m. med hjälp av högre utvecklade varelser för att försnabba evolutionen och styra dess förlopp. Teorierna om liv från yttre rymden (panspermia) kan emellertid inte förklara livets början i universum. Kännetecknande för levande varelser på jorden är: 1. Levande varelse har en organiserad byggnad, som i huvudsak grundar sig på organiska molekyler dvs molekyler som innehåller kol (C). 2. Levande varelser använder energi från omgivningen för att bygga upp nya molekyler. 3. Levande varelser uppehåller en komplicerad struktur och en inre balans (homeostasi). 4. Levande varelser växer och förökar sig och överför därmed sitt genmaterial till sin avkomma. 5. Levande varelser reagerar på retningar och förändringar i omgivningen. 6. Levande varelser har förmåga förändras - mutera - med hjälp av evolutionen. Virus uppfyller de flesta av de ovannämnda kriterierna. Virus kan emellertid inte föröka sig självständigt. De behöver alltid en värdcell. Av denna orsak anses virus inte vara levande varelser. Den prebiotiska världen Man tror att jordklotet bildades för 4,6 miljarder år sedan. De äldsta fossila fynden är över 3 miljarder år gamla. Den mellanliggande tiden - ca 1.5 miljarder år - anses vara av avgörande betydelse för livets uppkomst. Denna tidsperiod kallas prebiotisk tid (prebiotisk = före liv). Temperaturen på jorden var då över 70°C och både haven och regnen var sura. Den vulkaniska aktiviteten var hög och gav mycket värme. Solens strålar innehöll rikligt med UVstrålning på grund av att det absorberande ozonskiktet saknades. Åskväder var troligen mycket vanliga. Enligt den naturvetenskapliga teorin uppstod liv gradvis under en tid av hundratals miljoner år. De små organiska molekyler, som de levande varelserna använder som byggstenar, kan bildas utan levande varelser dvs. prebiotiskt. Polymeriseringen av de organiska molekylerna är av avgörande betydelse för uppkomsten av liv. En molekyl, som kan reglera uppkomsten av egna kopior, har överlägsna möjligheter jämfört med andra molekyler. RNA är en sådan molekyl och den anses vara bland de första polymererna. Antal atomer Molekylgrupp Under 25 Glukos, ammoniak, kväve, oorganiska salter Under 60 Aminosyror, nukleotider, socker, fettsyror Under 150 Fetter Över 1500 Proteiner, polynukleotider, polysackarider Över 100.000 Membraner, stödfibrer, proteoglykaner, nukleinsyror Molekylernas evolution Man vet tyvärr mycket lite om hur molekylernas evolution gick till. Ett är emellertid säkert, de organiska föreningar, som behövs för att skapa liv, skulle inte kunna bildas i dagens syrerika miljö. Man tror därför att den dåtida atmosfären var reducerande och innehöll vatten och väte samt bl.a. metan och ammoniak. Bilden visar Harold Ureys och Stanley Millers försöksanläggning, med vilken de på 1950-talet kunde visa att såvida man imiterar förhållandena på det "unga" jordklotet kommer enkla molekyler att reagera sinsemellan och bilda bl.a. aminosyror. 10% av kolet bildade små organiska föreningar och 4% bildade aminosyror. Samma typer av aminosyror påträffades år 1969 i en meteorit, som föll ner i Australien. Basen adenin (A), som förekommer i ATP och deltar i bindningen av energi samt i DNA, bildas vid ovanbeskrivna förhållanden. Även socker, kolväten och organiska syror bildas. Sähkövarauksia = elladningar Vesi = Vatten Lämpö = Värme Keruuallas - Uppsamlingskar RNA-katalys RNA innehar en central position vid molekylernas evolution. RNA har nämligen katalytiska egenskaper och därför möjlighet att reglera bildningen av egna kopior och andra RNA-molekyler. RNA har en struktur som möjliggör överförning av genmaterial till nästa generation. RNA förekommer huvudsakligen som enkelsträngat, vilket ger möjlighet till stor strukturell mångformighet. Man har under laboratorieförhållanden visat att en RNA-molekyl bestående av 52 nukleotider kan fungera som en katalysator. Man har vidare konstaterat att en enstaka katalytisk RNA- molekyl, ett sk ribotzym, kan vrida sig på två olika sätt, som har olika katalysiska egenskaper. Detta innebär att RNA-molekyler, som är enzymatiskt aktiva, kan ha ett gemensamt ursprung trots att de har olika strukturer och funktioner. RNA har en central roll vid proteinsyntesen i levande organismer. Budbärar-RNA (messenger-RNA = mRNA) bestämmer aminosyreordningen, transfer-RNA (tRNA) hämtar de rätta aminosyrorna i rätt ordning till den proteinproducerade enheten, ribosomen, där det ribosomala RNAet (rRNA) är av central betydelse. Flera typer av RNA-molekyler deltar alltså i proteinsyntesen. Ribotzymer RNA-molekyler med katalytisk aktivitet kallas RNA-enzymer eller ribozymer. Hit hör bl.a. vissa RNA introner med förmåga till "self-splicing" och endel viralt RNA. År 1981 visade Thomas Cech och hans forskargrupp att det encelliga urdjuret Tetrahymena thermophilus har förstadier till rRNA med förmåga att katalysera sin egen spjälkning till mogna rRNA-molekyler. År 1983 gjorde Sidney Altman liknande observationer. Han studerade ribonukleas P, som deltar i processeringen av tRNA-förstadier. Ribonukleas P är ett ovanligt enzym, som består av proteiner och RNA. Dess katalytiska aktivitet ligger i RNA-underenheten. Till skillnad från det självspjälkande rRNAet riktar sig den katalytiska aktiviteten hos ribonukleas P mot substrat utanför. Ribotzymerna anses vara en kvarleva från den s.k. RNA-världen. Ribotzymerna tros ha fungerat både som lager för biologisk information och som enzymer. Ribotzymernas katalytiska centrum består enbart av RNA. De behöver ingen proteindel vid katalysen. Den vanligaste ribotzymen i cellerna är ribosomen. Man har visat att ribosomerna i Thermus aquaticus bakterier, som lever vid mycket höga temperaturer, har ett katalytiskt centrum, som består nästan enbart av 23S rRNA. Urcellen På basis av över 3 miljarder år gamla fossila fynd tror man nu att de första cellerna liknade bakterier. De äldsta fossila fynden av kärnförsedda (eukaryota) celler är ca 1 miljard år gamla. På basis av evolutionsteorin framkastades tanken att allt liv härstammar från en enda urcell. I dag indelar man emellertid alla organismer i tre klasser: 1. bakterier 2. arkebakterier eller arkéer 3. kärnförsedda celler eller eukaryota celler. Arkéerna kan leva under mycket extrema förhållanden, såsom vid mycket höga temperaturer och salthalter. Man gjorde nyligen en jämförelse av gener i jäst, bakterier (9 arter) och arkéer (6 arter). Det visade sig att generna i jästens kärna liknar dem i arkéerna, medan generna i jästens cytoplasma påminner om dem i bakterierna. Detta tyder på att arkéerna i något skede kommit in i bakterierna och fortsatt att leva i dem som ett slags parasiter. Med tiden har arkéerna förlorat sina egna gener för reglering av ämnesomsättning. På samma sätt har skett med de gener som styr bakteriernas egen genreglering. På detta sätt anser man att den cell, som motsvarar dagens eukaryota cell, har uppstått. Arkit = arkéer Muut bakteerit = övriga bakterier Purppurabakteerit = purpurbakterier Syanobakterier = cyanobakterier Kloroplasti = kloroplast Kasvit = växter Eläimet = djur Sienet = svampar Mitokondrio = mitokondrier Muinainen aitotumallinen = tidig kärnförsedd cell = tidig eukaryotisk cell Arkit = arkéer Alkusolu = urcell Livets gener En fråga som länge sysselsatt forskare är "Hur många gener behövs för att en levande organism skall uppstå och bestå?" Mycoplasma genitalium har den minsta mängden gener. Dess genom består av ca 580 000 baspar och de bildar ca 470 gener. När man med hjälp av s.k. transposoner åstadkom slumpmässiga förändringar i M. genitaliums genmassa fann man att 265-350 av de 470 generna är nödvändiga för dess överlevnad. De övriga ca 120-205 generna kan man eliminera. Art Genomets storlek (miljoner Antal gener Antal letala mutationer baser) Mycoplasma genitalium 0.58 470 265-350 4.2 4100 Okänt 4.6 4288 ca 1880 13.5 25000 Okänt 135 25000 Okänt 180 13600 ca 3100 97 19099 Okänt 3330 32000 (?) Okänt Bacillus subtilis Escherichia coli (tarmbakterie) Saccharomyces cerevisae (jäst) Arabidopsis thaliana (backtrav) Drosophila melanogaster (bananfluga) Caenorhabditis elegans (nematod) Homo sapiens (människa) Cellens sammansättning Den molekylära sammansättningen och grundämnena i de levande organismerna skiljer sig avsevärt från den icke levande materians. Över 99% av de organiska molekylerna består av väte, kol, syre och kväve. Detta är grunden för begreppen organisk och oorganisk kemi. De biologiska molekylerna indelas i fyra grupper: 1. proteiner eller äggviteämnen 2. lipider eller fettämnen 3. kolhydrater 4. nukleinsyror Dessa molekyler har vissa gemensamma egenskaper. De byggs huvudsakligen upp av vissa grundenheter, som binds ihop av starka kovalenta bindningar, vilket ger upphov till stora makromolekyler. Processen kallas polymerisering. Proteinerna byggs upp av aminosyror, kolhydraterna av monosackarider och nukleinsyrorna av nukleotider. Lipiderna har inte en lika klar grundstruktur som de tre övriga men t.ex. fettsyror utgör ofta ett centralt element i dem. Kolhydraterna är de mångsidigaste biomolekylerna. Genom att slumpmässigt kombinera fyra olika nukleotider kan man åstadkomma 24 olika tetranukleotider medan man utgående från fyra monosackarider kan bilda 35560 olika strukturer. När det gäller oligosackaridernas struktur används dock vanligen en ganska begränsad uppsättning grundelement. Proteinerna Proteinerna är kväverika organiska föreningar, som är absolut nödvändiga för alla levande organismer. Proteinerna har många olika uppgifter i cellerna och deras kemiska egenskaper är mångsidiga. Proteiner är stora makromolekyler. Deras molekylvikt varierar mellan 30,000 och 500,000 Da. De är mångsidiga både till form och struktur. Proteinerna byggs upp av en eller flera molekylkedjor, s.k. polypeptider. Polypeptider byggs upp av aminosyror, som är kovalent bundna till varandra. I proteinerna, som kan bestå av hundratals aminosyror, förekommer endast 22 olika aminosyror. Dessa aminosyror kallas essentiella aminosyror. Vid sidan av dessa essentiella aminosyror används några andra aminosyror för andra ändamål i cellen. Lipiderna En stor del av cellens fetter består av fettsyror, som är bundna till en, två eller tre av glycerolets hydroxylgrupper (se bild). Det är fråga om mono-, di-, triacylglyceroler. En triacylglycerol består av en glycerol förestrad med tre fettsyror. Fosfolipiderna består av en glycerol, två fettsyror, en fosfatgrupp samt en polär alkohol. Alkoholgruppen är hydrofil. Glykolipider innehåller en eller flera sockermolekyler bl.a. D-glukos, D-galaktos eller N-acetyl-D-galaktosamin. Sockergrupperna är hydrofila. Steroiderna bildar en egen grupp fettämnen i cellen. Steroler innehåller en steroidkärna, som består av 4 ringar, till vilka ett varierande antal sidogrupper är bundna. Kolesterol är den viktigaste sterolen. Kolesterol förekommer i cellmembranen. Många hormoner är steroider. Sfingolipiderna är stora och ganska komplicerade molekyler, som förekommer bl.a. i cellmembranen. Klorofyll är också en lipid. Klorofyll består av en porfyrinring, till vilken en magnesiumjon (Mg) bundits. Klorofyll deltar i växternas fotosyntes. Karotenoiderna hör till samma grupp. Vaxer är glycerider, som består av långa, icke vattenlösliga fettsyror. De bildar ofta en fast fas. Vaxerna förekommer bl.a. på ytan av hud, hår och fjädrar. Terpener bildas från föreningar som kallas isoprener. De förekommer i vissa växtoljor och många av dem doftar. Till molekylerna, som ingår i cellernas signalsystem, fogas farnesyl eller isoprenyl. På detta sätt modifieras signalsystemets enzymer så att de blir fettlösliga, varefter de kan vandra ut i cellens membraner. De fettlösliga vitaminerna A, E och K är också terpener. Lipoproteiner bildas då fetter och proteiner går samman. Exempel på dessa är VLDL (Very Low Density Lipoprotein), LDL (Low Density Lipoprotein), och HDL (High Density Lipoprotein) som alla förekommer i blodet. Kolhydraterna Cellen innehåller många olika kolhydrater. Grundstrukturen för dem är monosackariderna, som syns på bilden nedan. Monosackariderna innehåller vanligen fem eller sex kolatomer (pentoser och hexoser). I vattenlösning antar socker en cyklisk form (se bilden). Hydroxylgruppens placering har en avgörande betydelse för monosackaridens kemiska egenskaper. Skillnaden mellan t.ex. glukosamin och galaktosamin (eller glukos och galaktos) beror på OH-gruppens orientering på kolatom fyra. (α-D-mannos; α-D-galaktos; α-L-fukos; α-D-N-acetylglukosamin; α-D-N-acetylgalaktosamin; N-acetylneuraminsyra (sialsyra)) Nukleinsyrorna Nukleinsyrorna är uppbyggda av nukleotider. En nukleotid innehåller en purin- eller pyrimidinbas, en ribos (RNA) eller deoxiribos (DNA) och en fosfatgrupp. I DNA förekommer fyra baser: cytosin (C), tymin (T), adenin (A) och guanin (G). I RNA har tymin ersatts av uracil (U). Cytosin, tymin och uracil är pyrimidiner medan guanin och adenin är puriner. Nukleotiderna binds till varandra via fosfatgrupperna och på så sätt bildas de mycket stora polymererna i DNA-molekylerna. Purinerna och pyrimidinerna binds till varandra med vätebindningar. Mellan A och T (eller U) bildas två vätebindningar och mellan C och G bildas tre. Den sistnämnda bindningen är den starkaste bindningen. Bilden visar hur DNA:s dubbelspiral är uppbyggd. Baserna är röda och sockerfostatdelarna blå. Bilden till vänster åskådliggör terminologin i samband med nukleotiderna. Baserna, med sina sockergrupper (ribos eller deoxiribos), kallas nukleosider. När en fostafgrupp binds till nukleosiden bildas en nukleotid. Bilden visar en adenosinmonofosfat. För nukleotiderna används följande förkortningar: AMP, CMP, UMP och GMP. (Adenin; Adenosin) RNA sönderfaller mycket lättare än DNA t.ex vid behandling med baser. I vår omgivning finns dessutom nästan överallt s.k. ribonukleaser, som enzymatiskt spjälkar RNA. Cellmembranet Cellerna omges av ett cellmembran, som består av ett dubbelt lipidskikt. Största delen av lipiderna hör till gruppen fosfolipider men cellmembranet innehåller också t. ex. kolesterol. I cellmembranet finns en stor mängd olika proteiner, t.ex. receptorer. Med hjälp av dessa membranproteiner fäster cellen vid andra celler och vid basalmembranen. Membranproteiner fäster också cellens inre strukturer. Cellmembranets proteiner har ofta sockergrupper kopplade till sig. Cellmembranet är ett semipermeabelt membran, vilket innebär att små molekyler, såsom t.ex. vatten, penetrerar membranet med hjälp av osmotiska krafter medan största delen av transporten genom cellmembranet regleras mycket noggrant. I cellmembranet förekommer speciella kanaler för t. ex olika joner eller socker. Dessa kanaler byggs upp av proteiner. På detta sätt kan cellen upprätthålla inre optimala betingelser, som klart avviker från dem i den yttre miljön. Även cellmembranets struktur är betydelsefull. Ett exempel på detta är mikrovilli på tunntarmens epitelceller, som förstorar ytarealen och därmed underlättar absorptionen av näringsämnen. Fosfolipider i dubbelmembranet; kolesterol; Transportprotein; Receptor Kärnan Kärnan är cellens största organell och den innehåller cellens genetiska material dvs DNA. Under cellernas delningsfas kondenseras DNA:et till kromosomer. Mellan delningarna (i interfas) är DNA:et lösare packat, i form av kromatin. Kärnan beskrevs för första gången år 1833 av Robert Brown. Kärna; Kärnmembran; Nukleol; Kromatin Kärnan är vanligtvis rund eller oval. I de neutrofila vita blodkropparna är kärnan loberad, dvs uppdelad i många olika delar. De flesta celler har en kärna. Däggdjurens mogna röda blodkroppar saknar emellertid kärna. Kärnan omges av ett kärnmembran, som har små öppningar, kärnporer. Via dessa kärnporer vandrar ämnen mellan kärnan och cytoplasman. Exempel på sådana ämnen är messenger-RNA, som är modell för proteinsyntesen i cytoplasman. Transporten mellan kärna och cytoplasma regleras av speciella transportprotein. Tidigare trodde man att kärnan var ganska "löst" organiserad. Nya undersökningar har emellertid visat att kärnan innehåller många olika specialområden. Vissa av dem är mycket dynamiska och utbyte av protein mellan dem sker snabbt. Cytoskelettet Cytoskelettet består av tre olika typer av fibrillära strukturer: 1. mikrotubuli 2. mikrofilament 3. intermediära filament. De olika filamenten bildar ett komplext nätverk, som fäster vid kärnan och cellmembranet. Cytoskelettet bestämmer cellens form. Mikrotubuli har dessutom till uppgift att transportera organeller och molekyler i cellen. Mikrofilamenten är mycket viktiga för cellens rörelser (bl.a. i muskelcellerna). Både mikrofilamenten och mikrotubuli sönderfaller och nybildas snabbt beroende på cellens olika behov. De intermediära filamenten är av mer bestående natur. Metabolism Cellens ämnesomsättning eller metabolism omfattar en oändlig mängd kemiska reaktioner. En av de viktigaste reaktionskedjorna är cellens energimetabolism. Den handlar om hur organismen skaffar sig närgingsämnen och den använder energin från dem för att upprätthålla livet. Cellens ämnesomsättning kan delas i två delar. 1. Katabolism = nedbrytningen av ämnen, som kommit utifrån eller lagrats i cellen. Härvid avges energi. 2. Anabolism = syntes av makromolekyler från enkla molekyler. Härvid krävs energi. En del av ämnesomsättningen sker i själva cytoplasman. Peroxisomer och mitokondrier deltar emellertid också i ämnesomsättningen och de effektiverar den. Vissa celler använder syre vid sin ämnesomsätttning (aerob metabolism), andra celler klarar sig utan syre (anaerob metabolism). Multicellulära organismer har många olika celltyper, som har olika uppgifter och olika metabolism. Extracellulär matrix Extracellulär matrix produceras av celler och finns runt alla celler. Det består av ett nätverk av proteiner och kolhydrater. I mångcelliga organismer fungerar extracellulär matrix både som stöd och som fästpunkter. Molekylerna i extracellulär matrix har olika fysikaliska egenskaper, vilket påverkar vävnadernas egenskaper såsom elasticitet och draghållfasthet. Extracellulär matrix kan vara mycket välorganiserat. Ett exempel på detta är basalmembranet under epitelcellerna. I bindväv, brosk och ben har extracellulär matrix mycket stor betydelse. Dessa vävnader består till största delen av extracellulär matrix. Molekylerna i extracellulär matrix kan indelas i tre huvudgrupper: 1. proteoglykaner 2. kollagen 3. adhesiva proteiner Proteoglykaner består av en central proteinkedja "core-protein", till vilken ett stort antal heteropolysackarider försedda med laddade kolhydrater är bundna. Kollagen ger stadga åt vävnaderna. De adhesiva proteinerna förankrar cellerna vid extracellulär matrix. Säkeinen kollageeni: kollagenfiber; Laminiini: laminin; Tenaskiini: tenascin; Aggregaani: aggrecan; Dekoriin: decorin; Hyaluronaani: hyaluronan; Fibronektiini: fibronektin; Tyypin IV kollageeni: typ IV kollagen; Perlekaani: perlecan Mikrober Med begreppet mikroorganismer eller mikrober förstår man enkla, vanligen encelliga, icke differentierade organismer. De är icke synliga med blotta ögat. Till mikroberna räknas bakterier, alger, urdjur, jäst, mögel och virus. Mikrober förekommer överallt i vår miljö. En del av mikroberna förorsakar sjukdomar andra är nyttiga t.o.m. livsviktiga för oss. Mikroberna bryter ner död materia i naturen. Detta är mycket viktigt. I läkemedelsindustrin och livsmedelsindustrin används många olika mikrober. Sienet: Svampar; Alkuelämimet: Urdjur; Virukset:Virus; Home: Mögel; Hiiva: Jäst; Jäkälät ja mykoritsat: Lavar och mykoritsor; Bakteerit: Bakterier Signalering Cellen registrerar hela tiden sin miljö och de förändringar, som sker där. Förändringarna registreras som retningar, som vid en viss nivå åstadkommer en aktivering av det inre cellulära retningsledningsystemet. Cellsignaleringen börjar vid receptorn. Under signaleringrutten förstärks signalen och responsen kan synas t.ex. som förändringar i cellens ämnesomsättning eller i genernas expression. Att märka är att cellen hela tiden utsätts för många olika retningar, som inte enskilt ger upphov till förändringar i cellens funktioner. Retningar som sker samtidigt kan dessutom uppväga varandras responser och därmed fungera som ett slags buffertar. En respons uppstår endast då tröskelvärdet överskrids. Ligandi/Ärsyke: Ligand/retning; Resprori; Receptor; Viestintävälittysreitti: Signaleringsrutt; T Tilannekohtainen viestintävälitysreitin muokkaus: Modulation "på vägen" ; Ärsykkeen vahvistaminen: Förstärkning av retningen; Aineenvaihdunnan säätely: Reglering av ämnesomsättning; Geenien ilmentymisen säätely: Reglering av genexpression; Solun tukirangan muutokset: Förändringar i cellens stödskelett. Tillväxt En levande organism kan tillväxa på två olika sätt: 1. genom att de enskilda cellerna ökar i storlek. Detta kallas hypertrofiering. 2. genom att antalet celler ökar vilket kallas proliferation. När det gäller encelliga organismer är tillgången på näring den avgörande faktorn för tillväxthastigheten. Sålunda kan E. coli vid optimala betingelser dela sig en gång var 20.-30. minut. I mångcelliga organismer förekommer olika cellpopulationer. Vissa celler delar sig hela tiden, andra delar sig nu och då, en del celler delar sig t.ex. som en följd av en skada. Vissa celler delar sig aldrig. En vuxen människa måste dagligen producera många miljoner nya celler för att ersätta de som nöts bort och de som dött genom programmerad celldöd (apoptos). Ju lägre differentieringsnivå en cell befinner sig på desto bättre delningsförmåga har den. Förökning Cellerna kan dela sig på två sätt, antingen mitotiskt eller meiotiskt. Mitosen är en könlös celldelning och genomet överförs oförändrat till dottercellerna. Under meiosen förändras och omfördelas genomet i samband med att spermier och äggceller bildas. Alla organismer med könlig förökning har meiotisk delning. Hos honor bildas endast en äggcell och tre förkrympta polkroppar. Hos hanar bildas fyra mogna spermier. EM bild av tjurspermier i längdsnitt (Pitkittäisleikkaus) och tvärsnitt (Poikkileikkaus). I tvärsnitten ses svansens struktur. Cellens delning För att en mångcellulär organism skall tillväxa behövs självfallet celldelning. Under den tidiga embryonalutvecklingen är celldelningen mycket livlig. Sedan följer en tid av mognad och celldifferentiering. Celldelning behövs emellertid hela tiden enär kroppens celler har en begränsad livslängd och måste därför ersättas. En celldelning medför alltid en risk för den enskilda cellen. För att minimera risken har en komplicerad kontrollmekanism utvecklats. Under cellcykeln finns så kallade "kontrollstationer", vid vilka cellen försöker säkerställa att delningen framskridit på rätt sätt. I denna kontroll deltar ett stort nätverk bestående av proteinkinaser, specifika kinasinhibitorer, transkriptionsfaktorer och specifika proteaser. När förhållandena inne i och utanför cellen är lämpliga för delning lossnar specifika proteinkinaser från sina regulatorer. Härvid inverkar vissa nyckelproteiner. Detta utlöser en kedjereaktion, som leder till att DNA fördubblas och att cellen delar sig. Celldöd En cell kan dö på två sätt: 1. genom apotos 2. genom nekros. Apoptos kallas ofta programmerad celldöd och innebär att cellen gör självmord. Apoptos är en naturlig del av organismens normala liv. Nekros innebär att cellen dör på ett okontrollerat sätt eller genom en olycka. Nekros omfattar ofta många celler medan apoptos gäller en enskild cell i gången. Nekros leder till inflammation. Vid apoptos uppkommer ingen inflammation. Bilden visar Apoptos till vänster och Nekros till höger Forskning (Cellbiologi) Man studerar celler och deras funktioner med många olika metoder. De mikroskopiska metoderna har en i nyckelställning då man undersöker cellulära responser. Med ljusmikroskop (LM) är det förhållandevis enkelt att studera celler, också levande celler. Med elektronmikroskop (EM) studerar man cellorganeller och cellens inre struktur. Tekniken för EM är betydligt mer krävande än den som behövs för LM. På den molekylära nivån finns en hel del biokemiska metoder, med vilka man studerar cellens molekylära struktur och molekylernas växelverkningar. Dessa metoder grundar sig vanligtvis på att de molekylerna, som man är intresserade av, isoleras från cellens andra molekyler. Detta innebär naturligtvis att cellerna eller vävnaderna måste sönderdelas. Med molekylärbiologiska metoder är det möjligt att studera genernas funktioner och deras reglering. Man kan överföra specifika gener till celler och på så sätt studera genmutationernas inverkan på cellens funktioner. Med molekylärbiologiska metoder är det möjligt att framställa märkmolekyler, som kan fogas till gener som kodar för speciella proteiner. Med denna metod kan man följa enskilda molekylers placering i levande celler i realtid. Härvid behövs ett fluorescensmikroskop. Cytokemiska metoder Ljus- och elektronmikroskopiska metoder kan användas för att lokalisera olika molekyler och den metaboliska aktiviteten inne i celler. Härvid kombinerar man olika cytokemiska metoder, som bygger på antikroppar, enzymer och radioaktiva föreningar. Cellodling Med cellodlingsteknik kan man studera celler och deras funktioner under noggrant kontrollerade förhållanden. Olika djur-, växt- och bakterieceller kan odlas in vitro i laboratorier. För att växa i cellkulturer behöver cellerna kontrollerade yttre betingelser vad det gäller näringsämnen, temperatur och pH-värden. Celler kan odlas i suspension eller på fast underlag. I suspension är det möjligt att odla celler i stora kvantiteter. Många djurceller kan emellertid inte odlas i suspension. Dessa celler måste odlas på täckglas eller plastytor i flaskor eller skålar med näringslösning. I dem bildar cellerna ett s.k. monolayer dvs ett enskiktat lager av celler. Bakterier kan odlas i suspension eller på ytan av ett halvfast medium. Det halvfasta mediet är kolhydratrikt och består av agar med tillsats av näringsämnen. Den specifika bakterien inokuleras i odlingsskålen. De enskilda bakterierna bildar kolonier i skålen. Cell- och molekylärbiologi: historik Andreas Vesalius (1514-1564) anses vara den moderna anatomins grundare. Han arrangerade de fösta offentliga dissektionerna i den anatomiska teatern i Padua. År 1543 publicerade han "De Humani Corporis Fabrica", som beskriver människans anatomi. Före Vesalius hade man studerat människokroppen på många olika, men rätt osystematiska sätt. Termen cell användes för första gången år 1665 då Robert Hooke (1635-1703) presenterade sina observationer över korkträdets uppbyggnad (se bilden nedan). Våra insikter i cellbiologi har alltså ackumulerats under en ganska kort tid dvs ca 350 år. Parallellt med att mikroskopen utvecklades förbättrades vår uppfattning om cellerna. Cellbiologins utveckling var ganska långsam i början. Kännedom om cellens funktioner och uppbyggnad ökade först under 1800-talet. Cellernas biokemiska reaktioner kartlades och deras organeller beskrevs. Detta ledde till att en cellteori formulerades. Under 1900-talet har cellbiologin utvecklats med en svindlande fart. Molekylärbiologin, som bygger på kännedomen om DNA molekylens byggnad och funktioner, har haft en stor betydelse. Nya cellbiologiska metoder har lett till att nya områden inom cellbiologin har utvecklats. I dag kan man följa med cellernas och deras proteiners funktioner i realtid med hjälp av fluorescerande fusionsproteiner. Under 1900-talet har cellbiologin utvecklats med svindlande fart. Molekylärbiologin, som bygger på kännedom om DNA molekylens byggnad och funtioner, har haft en stor betydelse. Nya cellbiologiska metoder har lett till att nya områden inom cellbiologin har utvecklats. I dag kan man följa med cellernas och deras proteiners funktioner i realtid med hjälp av flourescerande fusionsproteiner. 1600-talet Galileo Galilei (1564-1642) och Anthony Leeuwenhoek (1632-1723) anses vanligen vara de första som konstruerade ljusmikroskop. Sannolikt var det emellertid Zacharias Jansen (1580-1632) eller möjligen hans far som byggde det första ljusmikroskopet 1595. På 1600-talet trodde man allmänt att levern bildar blod utgående från maten, som man ätit. Man trodde vidare att organismen använder blodet som energikälla, dvs som en form av bränsle. År 1628 publicerade William Harvey (1578-1657) verket "De Motu Cordis et Sanguinis", i vilket han noggrant beskriver blodkretsloppet - både det lilla krestloppet från hjärtats högra kammare till lungorna och det stora kretsloppet från hjärtats vänstra kammare via aortan ut i kroppen. Man känner till endast ett fåtal publikationer över mikroskopiska iakttagelser från 1600-talet. Detta trots att Galileo Galilei hade tillgång till mikroskop från och med år 1607. Marcello Malphighi (1628-1694) utgav den första betydande publikationen i anatomi och cytologi. År 1660 beskrev han blodkretsloppet i organismen och bevisade därmed Harveys upptäckter. Malphighi beskrev kapillärnätet i grodornas lungor och de röda blodkropparna. Han anses vara utvecklingsbiologins och histologins fader. Hans namn återfinns i de Malphigiska kropparna i njurarna. Termen "cell" presenterades för första gången av engelsmannen Robert Hooke (1635-1703). Han undersökte korkträdet med mikroskop och fann en massa tomma hålrum, som han kallade "celler". Robert Hooke var i likhet med sin förman Robert Boyle intresserad av fysik och utvecklade Hookes lag. Tillsammans med Boyle studerade Hooke olika gaser. Hooke och Isaac Newton var ofta av olika åsikt. Mellan åren 1660-1690 byggde Anthony van Leeuwenhook (1632-1723) många egna mikroskop (ca 500 stycken). Han beskrev levande celler, bakterier, urdjur, spermier och röda blodkroppar. Leeuwenhoek hade ingen formell vetenskaplig skolning utan var närmast en skicklig konstruktör. Från och med år 1673 skrev han om sina upptäckter till Royal Society of London, som översatte dem till engelska och latin och publicerade dem i sin egen tidning. 1700-talet Marie Francis Xavier Bichat (1771-1802) dog ung men han anses i alla fall vara mycket viktig för utvecklingen av den moderna histologin och patologin. Han var den första som införde begreppet vävnad och han insåg sambandet mellan många sjukdomar och förändringar i vävnaderna. Bichat gjorde alla sina observationer i samband med dissektioner. Han använde inte mikroskop. Lazzaro Spallanzani (1729-1799) visade att det behövs både äggcell och spermie för att en ny individ skall bildas. År 1780 publicerade han sina första resultat av konstbefruktning på hundar. På den tiden trodde man allmänt att levande organismer uppstår ur död materia. Fenomenet kallas generatio spontana. Spallanzani visade att mikroorganismer dör vid kokning. Han var bland de första som motsatte sig generatio spontana. Fransisco Redi hade visserligen redan 1668 visat att flugor inte uppstår ur ruttnande kött, ifall man hindrar dem att lägga ägg i köttet. Giovanni Baptista Morgagni (1682-1771) var professor vid universitetet i Padua ända till sin död och är av central betydelse för utvecklingen av patologin. År 1761 publicerade han en sammanställning över ca 700 patienter. Han insåg att tuberkulos är en smittsam sjukdom, vilket ledde till att man började desinficera de rum, som tuberkulospatienter dött i samt bränna deras kläder. Han var mycket intresserad av fysiologi och beskrev bland annat symtomen vid angina pectoris. På 1700-talet var kemin en mycket outvecklad vetenskapsgren. Antoine Laurent Lavoisier (1743-1794) var den kemist, som upptäckte att grundämnen reagerar med varandra och bildar nya kemiska föreningar. År 1789 publicerade han verket "Traité élémentaire de chimi", som är den första moderna läroboken i kemi. Han visade att andning förbrukar syre och producerar koldioxid. Under den franska revolutionen fängslades han och år 1794 dömdes han till döden genom giljotinering. Han anklagades för att ha deltagit i böndernas revolt. När han bad om nåd för att få slutföra sina vetenskapliga experiment svarade domaren "Republiken Frankrike behöver inga vetenskapsmän". I Indien och Kina kände man redan för ca 3000 år sedan till att man kan skydda sig för den dödliga smittkoppsinfektionen genom att vaccinera med kokoppor (vaccinia). Engelsmannen Edward Jenner (17491823) påvisade experimentellt att vaccinering med kokoppor skyddar mot smittkoppor. 1800-1850 Ännu i början av 1800-talet härskade uppfattning om att liv uppehålls av s.k. vitala krafter (vitalism). Man trodde då att det var omöjligt att syntetisera organiska föreningar utgående från oorganiska grundämnen. Friedrich Wöhler (1800-1882) kunde år 1828 kullkasta denna uppfattning, då han syntetiserade urinämne utgående från ammoniumcyanat. Han var också den första som isolerade beryllium och aluminium. På 1820-talet studerade Wöhler i Stockholm hos Berzelius och de publicerade tillsammans en första teorin om kemisk katalys. De framkastade att jäsning är en katalytisk reaktion. Robert Brown (1773-1858) var en skotsk botaniker, som år 1801 gjorde en expedition till Australien. Han hämtade tusentals, för den västerländska världen okända, växter med sig hem. Klassificeringen av detta material tog över 5 år. År 1831 gav han namn åt cellens kärna. Då Charles Darwin (1809-1882) förberedde sin expedition till Amerika med HMS Beagle fick han undervisning i växtinsamlandets konst av Robert Brown. När Darwin kom hem behöll han emellertid själv sitt växtmaterial. Han ansåg att Brown hade tillräckligt att göra med sitt eget material. År 1827 visade Brown att mikroskopiska partiklar rör sig slumpmässigt då de blandas i en vätska. Detta kallas Browns rörelse. Matthias Schleiden (1804-1881), som först verkat som advokat, visade år 1838 att växter byggs upp av celler. Han visade också att det är vattenmolekylen och inte koldioxidmolekylen som sönderdelas vid fotosyntesen. Han skrev i mitten av 1800-talet en grundlig lärobok i botanik. Fotosyntesen hade beskrivits redan 1804 av Nicholas-Theodore de Saussure. Theodore Schwann (1810-1882) visade år 1839 att också djurens kroppar består av celler. Samma år beskrev Hugo von Mohl en normal celldelning, en mitos. Detta fullbordade cellteorin. Theodore Schwanns doktorsavhandling behandlade hönsäggets utveckling och speciellt hönsembryots andning. Dessutom var han intresserad av ämnesomsättningen och näringsämnenas spjälkning. År 1836 visade han att magens pepsin har förmåga att spjälka äggviteämnen i födan. Schwann var den första som beskrev nervtrådarnas myelinskida. Därav namnet Schwanns celler. 1900-1950 År 1921 isolerade kanadensaren Frederick Grant Banting (1981-1941) insulin ur bukspottkörtlar från hund. Under de följande åren utvecklade han en metod att vårda sockersjuka patienter. Orsaken till Bantings intresse för medicin och i synnerhet sockersjuka berodde på att en av hans barndomsvänner avled till följd av denna sjukdom. Banting blev professor vid det nya Western University of Ontario. Universitetet saknade emellertid lämpliga laboratorier, varpå Banting vände sig till professor John James Richard Macleod (1876-1935), som var professor i fysiologi vid universitetet i Toronto och bad om lov få att använda laboratorierna där. Banting arbetade därefter vid universitetet i Toroto och som assistent hade han med. stud. Charles H. Best (1899-1978). 10 hundar användes som försöksdjur. Banting och Best påvisade att extrakt från bukspottkörtlar är verksamt vid vården av sockersjuka. Detta ledde till att Macleod beslöt att ändra hela forskningsinriktningen vid sitt laboratorium till studiet av insulin. Biokemisten James Bertram Collip (1892-1965) hade stor betydelse vid utvecklingen av produktion av insulin i industriell skala. År 1923 utdelade Nobelkommittén Nobelpriset i fysiologi och medicin till Banting och Macleod. Man förvånades allmänt över att inte Best också fick del av priset och Banting tog emot sitt pris först efter noggrant övervägande. Han delade sedan priset med Best. Enligt Macleod hade även Collip varit värt priset och han delade därför sina prispengar med honom. Skotten Alexander Fleming (1881-1955) blev världsberömd när han kunde påvisa att penicillin har en bakteriedödade verkan. Som ung bodde Fleming lika nära tre olika medicinska universitet. Han valde St. Marys universitet för att han hade spelat vattenpolo med studerande från detta universitet. Han beslöt att specialisera sig i kirurgi men det hade inneburit att han borde ha flyttat till ett annat universitet. Fleming var emellertid en utmärkt skytt och övertalades av skytteklubbens kapten att stanna kvar vid St Marys. Han förblev detta universitet trogen under hela sitt liv. Han arbetade vid St Marys vaccinationscentrum. År 1928 upptäckte han en mögelhärd i en av sina bakteriekulturer och såg att bakterierna runt mögelhärden hade dött. Följande år publicerade han detta men det väckte då ännu ingen särskild uppmärksamhet. Under andra världskriget märkte man att penicillin var mycket effektivt mot bakterieinfektioner. År 1945 fick Alexander Fleming Nobelpriset i medicin tillsammans med Howard Walter Florey (1898-1968) och Ernst Boris Chain (1906-1979). Hans Adolf Krebs (1900-1981) undersökte bl.a. leverns syntes av urinämne, urinsyra och purinbaser samt dessutom mitokondriernas roll vid cellandningen. Krebs beskrev citronsyracykeln i mitokondrierna. Den kallas allmänt Krebs cykel. Krebs fick Nobelpriset i medicin år 1953. Elektronmikroskopet innebar ett mycket stort framsteg för den cellbiologiska forskningen. Ernst Ruska (1906-1988) och Max Knoll konstruerade det första elektronmikroskopet år 1931 i Tyskland. Ruska utvecklade mikroskopet vidare. År 1939 levererades det första elektronmikroskopet av Siemens. År 1986 fick Ernst Ruska Nobelpriset i fysik. 1950-2001 I början av sin långa karriär studerade Linus Pauling (1901-1994) de kemiska bindningarnas natur och han utvecklade regler för bildningen av kovalenta bindningar. Senare undersökte han bl.a. proteinernas struktur. Han påvisade att det sker strukturella förändringar i hemoglobinmolekylen då syre binds. Senare förklarade han bakgrunden till sicklecellanemi. År 1950 publicerade han en banbrytande artikel om proteinernas struktur. Denna upptäckt och hans tidigare forskningsresultat ledde till att han fick Nobelpriset i kemi år 1954. Pauling var en fredskämpe och protesterade kraftigt mot utvecklingen av kärnvapen. Detta ledde till att han tilldelades Nobels fredspris 1962. Linus Pauling är hittills den enda som fått Nobelpris från två olika områden. Det är mycket svårt att studera cellens organeller med enbart ljusmikroskop. Utvecklingen av elektronmikroskopi och cellfraktionering på 1950- och 1960- talet ledde till att man kunde studera cellen och dess funktioner på ett mycket bättre sätt. George Palade och hans forskargrupp har en central roll i detta sammanhang. De beskrev bl.a. ribosomerna, exocytosen och transporten av membranmaterial. Han utvecklade tekniken för framställningen av tunnsnitt för elektronmikroskop. År 1974 fick George Palade Nobelpriset i medicin. I Finland är hjärt- och kärlsjukdomarna ett stort folkhälsoproblem. På 1960-talet blev det klart att kolesterol har stor betydelse vid utvecklingen av dessa sjukdomar. År 1965 fick Konrad E. Bloch Nobelpriset i medicin för sina studier av kolesterolsyntesen. År 1985 fick Michael S. Brown och Joseph L. Goldstein Nobelpriset i medicin för sina studier av regleringen av kolesterolets ämnesomsättning. Cellens proteiner behövs på många olika ställen. Proteinerna identifieras ofta med hjälp av en mängd korta, sinsemellan olika aminosyrasekvenser och distribueras därefter till rätta platser. Günter Blobel och hans forskargrupp är av central betydelse i detta sammanhang. Blobel fick Nobelpris i medicin 1999 för sina insatser på detta område. Utvecklingen av molekylärbiologin Termen molekylärbiologi användes för första gången år 1938 av Warren Weaver (1894-1978). I vidare bemärkelse omfattar termen alla studier av makromolekylerna, deras funktioner och reglering. Nuförtiden används termen främst i samband med studier av DNA och RNA molekylerna och deras funktioner. År 1871 publicerade Johann Friedrich Miescher (1844-1895) en artikel, i vilken han beskriver DNA, som han isolerat från vita blodkroppar. Han använde termen nuklein. Detta ämne har en ovanlig sammansättning i och med att det innehåller rikligt med fosfor. Dessutom är det fråga om en syra. År 1874 beskrev han ett s.k. protamin, som han isolerat från laxrom. År 1889 visade Richard Altmann (1853-1927) att nuklein innehåller vissa sockergrupper och kväverika baser och att de bildar stora makromolekyler. Altmann använde beteckningen nukleinsyra. År 1929 visade Phebus Levene att sockerarten är deoxiribos. DNAet packas tätt i cellkärnan med hjälp av basiska proteiner s.k. histoner. Histonerna beskrevs av Albrecht Kossel redan år 1884. Gregor Mendel (1822-1884) studerade nedärvningen av synliga egenskaper dvs. färger och former i ärtväxter i Brno. Han såg vissa tydliga regelbundenheter. År 1865 publicerade han sina resultat men de väckte då ingen större uppmärksamhet. År 1900 hänvisar Hugo de Vries, Carl Correns och Ernst von Tschermar alla tre oberoende av varandra till Mendels resultat. 1900-1950 talet (Molekylärbiologin) Walter Sutton (1877-1916) (till höger), William Cannon och Edward Wilson kunde med stöd av mikroskopiska observationer föreslå att generna ligger i kromosomerna. År 1902 presenterade Sutton en kromosomteori för ärflighet. Han myntade termen gen. Han upptäckte kromosomernas rörelser vid den meiotiska celldelningen och år 1903 publicerade han tillsammans med Theodor Boveri en artikel, som bevisar att köncellerna är haploida dvs. att de innehåller bara en uppsättning av varje kromosom. Thomas Hunt Morgan (1866-1945) (till vänster) visade år 1907 att kromosomerna är av central betydelse vid överföringen av arvsmassa från en generation till den följande. Morgan studerade bananflugor och påvisade förekomsten av mutationer. År 1910 påvisade han förekomsten av könskromosomer i bananflugor. Han konstaterade att ögonfärgen är könsbunden. År 1926 publicerade han genteorin, som till en stor del bygger på George Mendels lagar. Morgan fick Nobelpris i medicin år 1933. George Beadle och Edward Tatum visade att generna styr bildningen av enzymer i cellerna och att mutationer åstadkommer förändringar i enzymernas byggnad och ofta också deras funktioner. Fastän deras teori "en gen-ett enzym" inte helt stämmer, skapade de grunden till den moderna uppfattningen om gener. År 1958 fick Beadle och Tatum tillsammans med Joshua Lederberg Nobelpriset. Oswald Avery , Colin McLeod och Maclyn McCarty påvisade att den ärftliga faktorn som överförs från en bakterie till nästa är DNA. Martha Chase och Alfred Hershey bekräftade dessa rön genom att använda protein- och DNA-prov infekterade med radioaktivt märkta bakteriofager (bakteriofag = virus som infekterar bakterier).